

Multimodal integration of carbon dioxide and other sensory cues drives mosquito attraction to humans
- PMID: 24581501
- PMCID: PMC4007582
- DOI: 10.1016/j.cell.2013.12.044
Multimodal integration of carbon dioxide and other sensory cues drives mosquito attraction to humans
Abstract
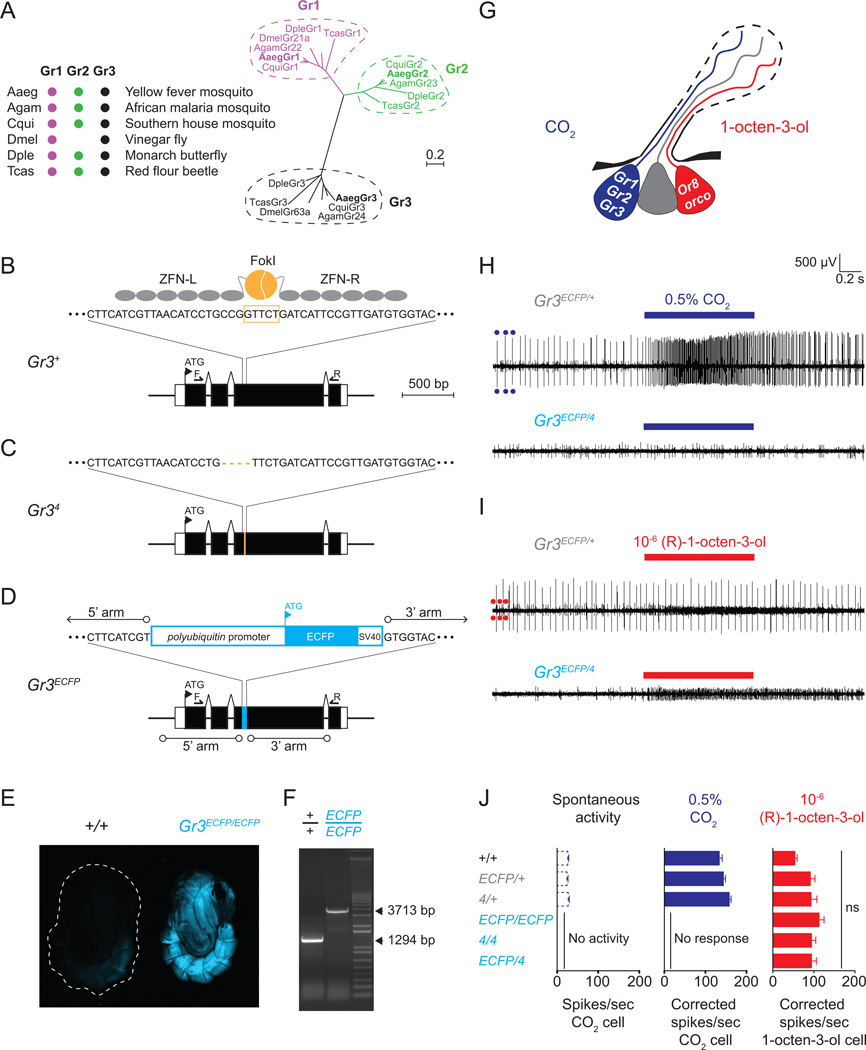
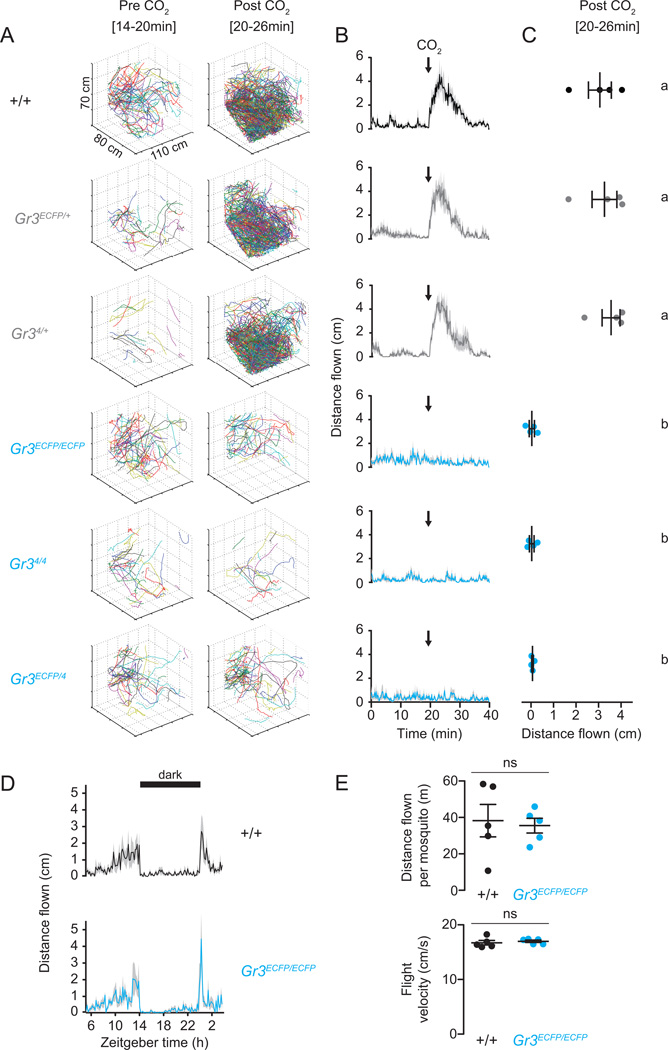
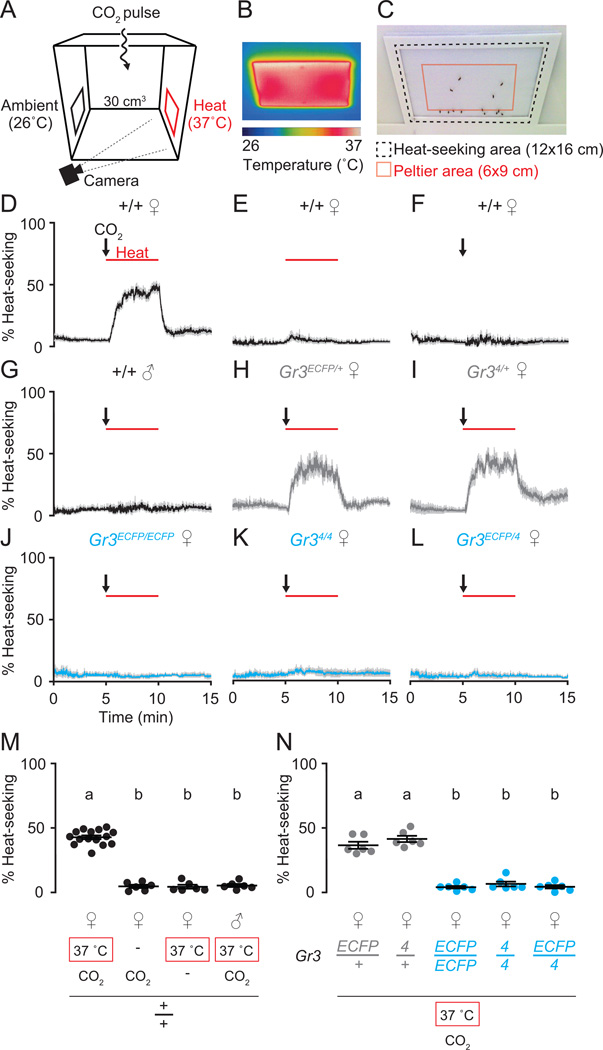
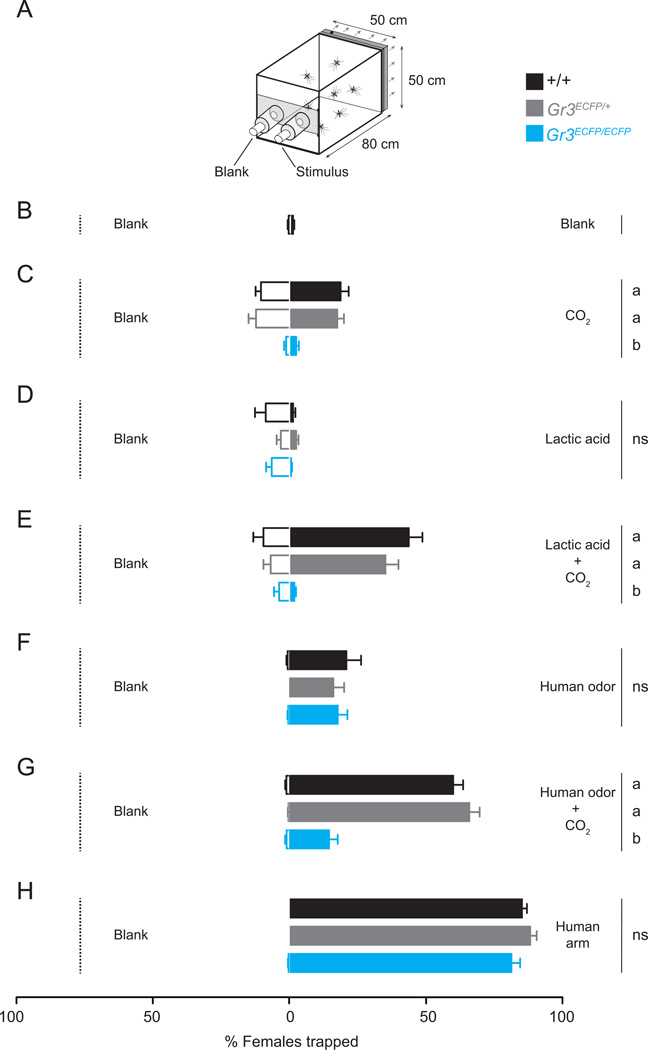
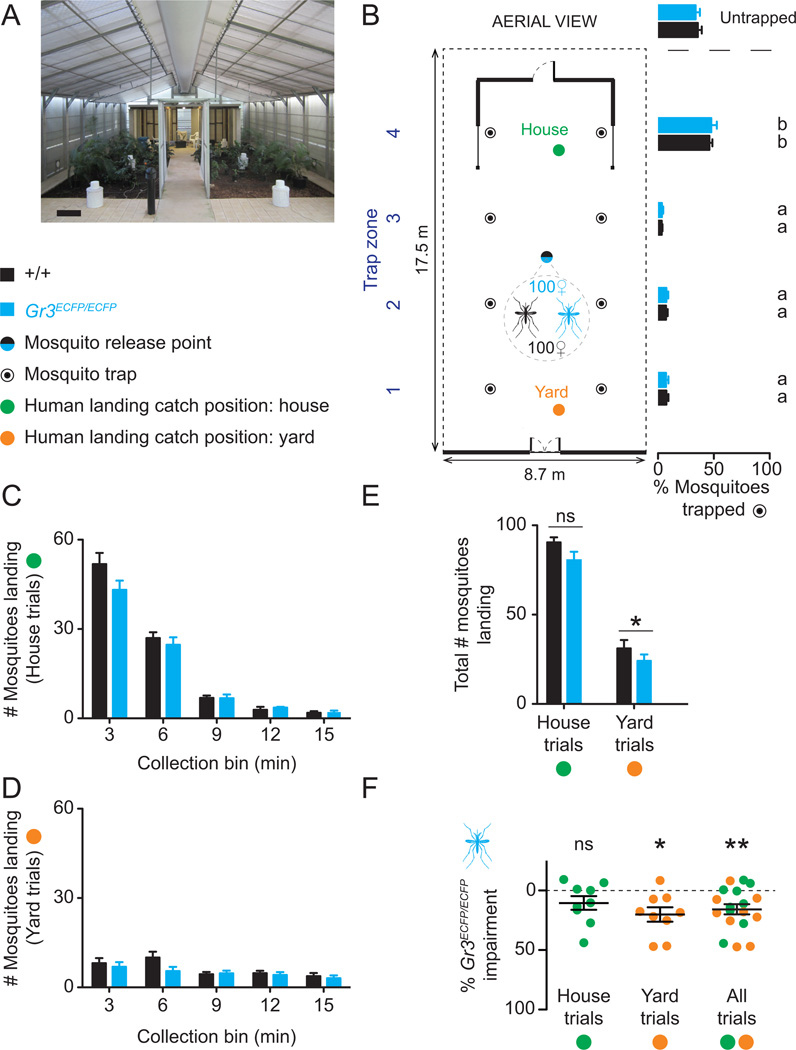
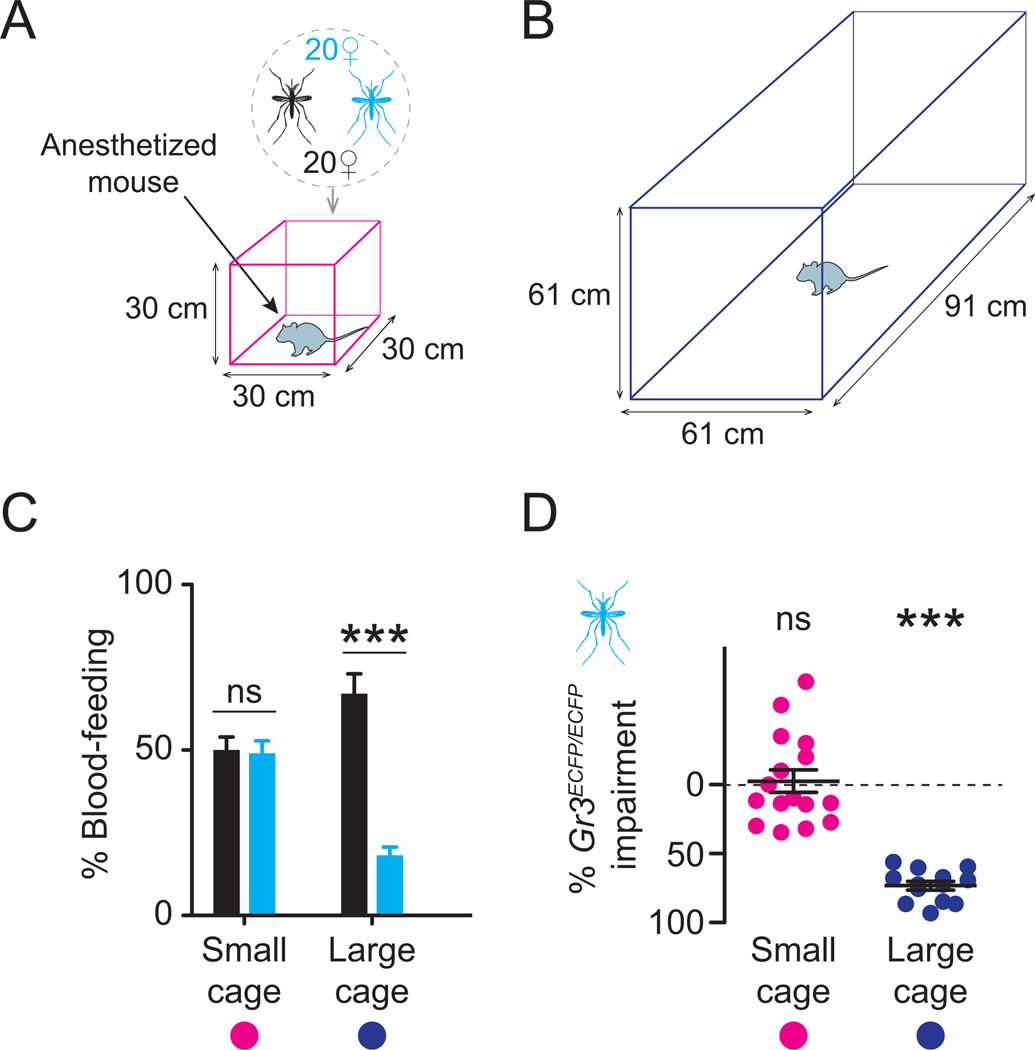
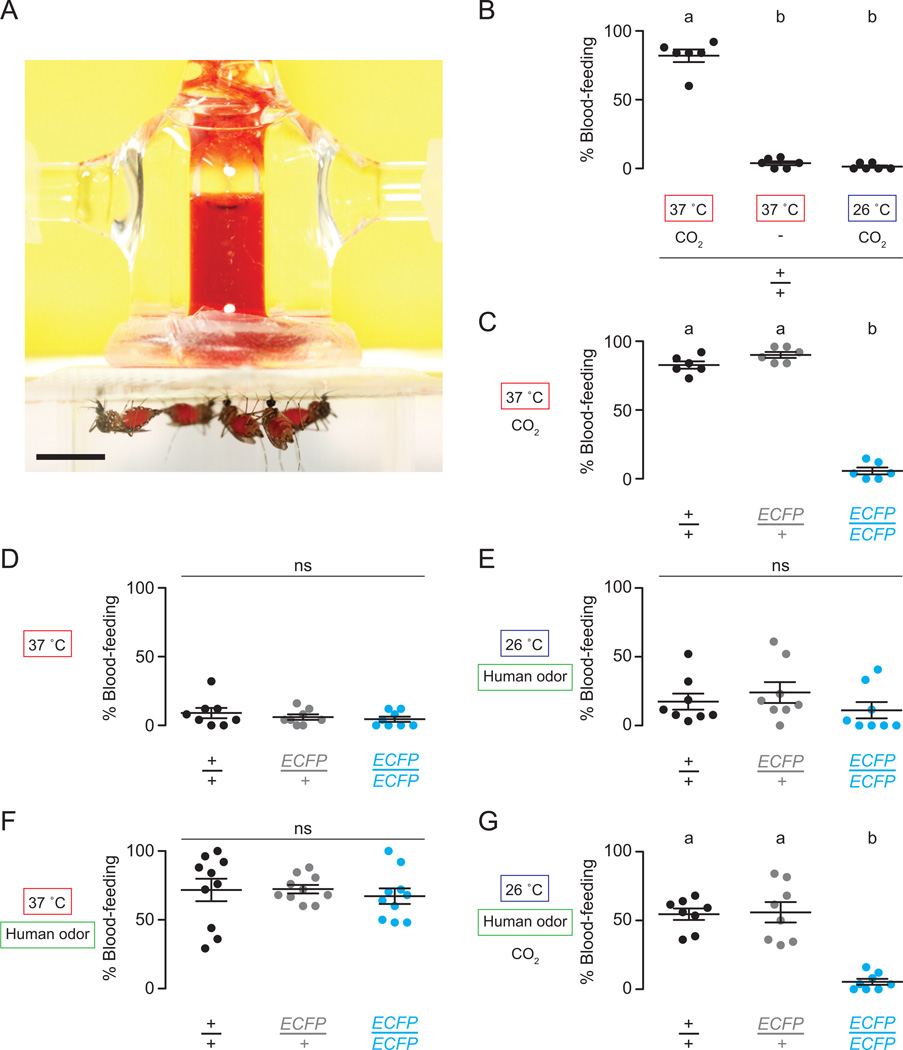
Multiple sensory cues emanating from humans are thought to guide blood-feeding female mosquitoes to a host. To determine the relative contribution of carbon dioxide (CO2) detection to mosquito host-seeking behavior, we mutated the AaegGr3 gene, a subunit of the heteromeric CO2 receptor in Aedes aegypti mosquitoes. Gr3 mutants lack electrophysiological and behavioral responses to CO2. These mutants also fail to show CO2-evoked responses to heat and lactic acid, a human-derived attractant, suggesting that CO2 can gate responses to other sensory stimuli. Whereas attraction of Gr3 mutants to live humans in a large semi-field environment was only slightly impaired, responses to an animal host were greatly reduced in a spatial-scale-dependent manner. Synergistic integration of heat and odor cues likely drive host-seeking behavior in the absence of CO2 detection. We reveal a networked series of interactions by which multimodal integration of CO2, human odor, and heat orchestrates mosquito attraction to humans.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Acree F, Jr, Turner RB, Gouck HK, Beroza M, Smith N. L-Lactic acid: a mosquito attractant isolated from humans. Science. 1968;161:1346–1347. - PubMed
-
- Anton S, van Loon JJ, Meijerink J, Smid HM, Takken W, Rospars JP. Central projections of olfactory receptor neurons from single antennal and palpal sensilla in mosquitoes. Arthropod Struct Dev. 2003;32:319–327. - PubMed
-
- Burgess L. Probing behavior of Aedes aegypti (L.) in response to heat and moisture. Nature. 1959;184:1968–1969.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
