

EFCAB7 and IQCE regulate hedgehog signaling by tethering the EVC-EVC2 complex to the base of primary cilia
- PMID: 24582806
- PMCID: PMC4027042
- DOI: 10.1016/j.devcel.2014.01.021
EFCAB7 and IQCE regulate hedgehog signaling by tethering the EVC-EVC2 complex to the base of primary cilia
Abstract
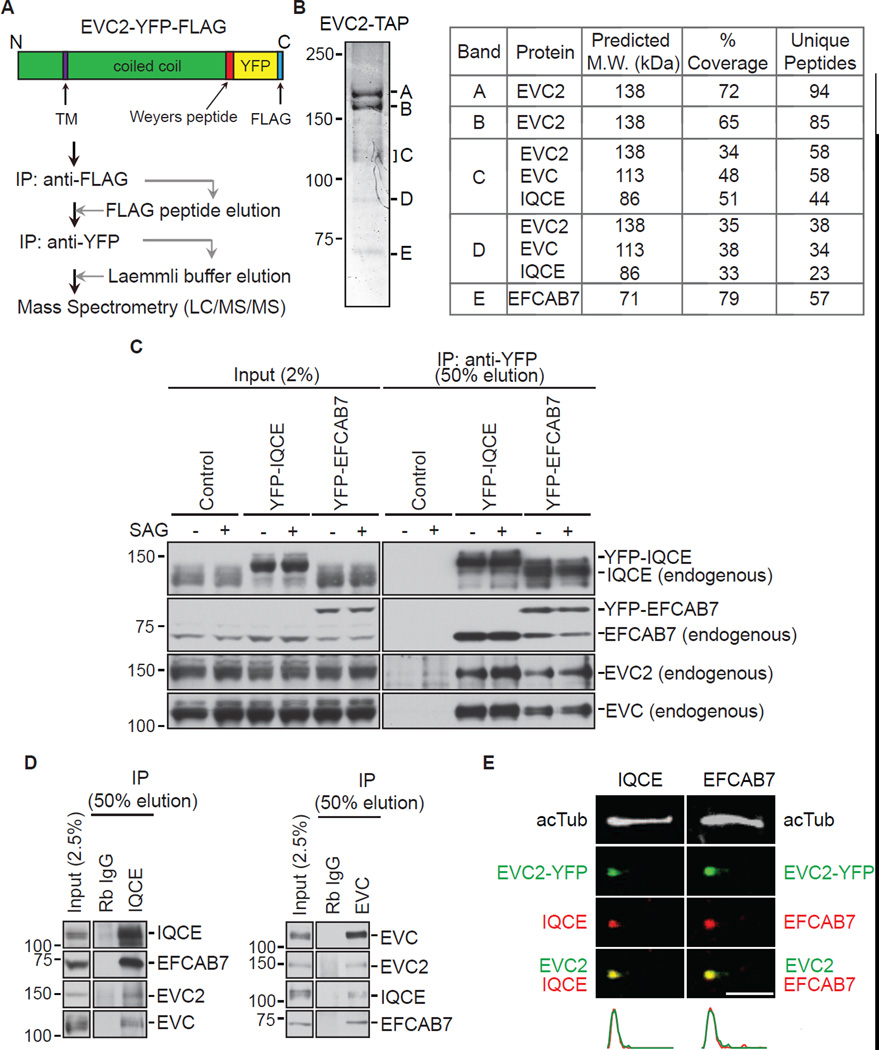
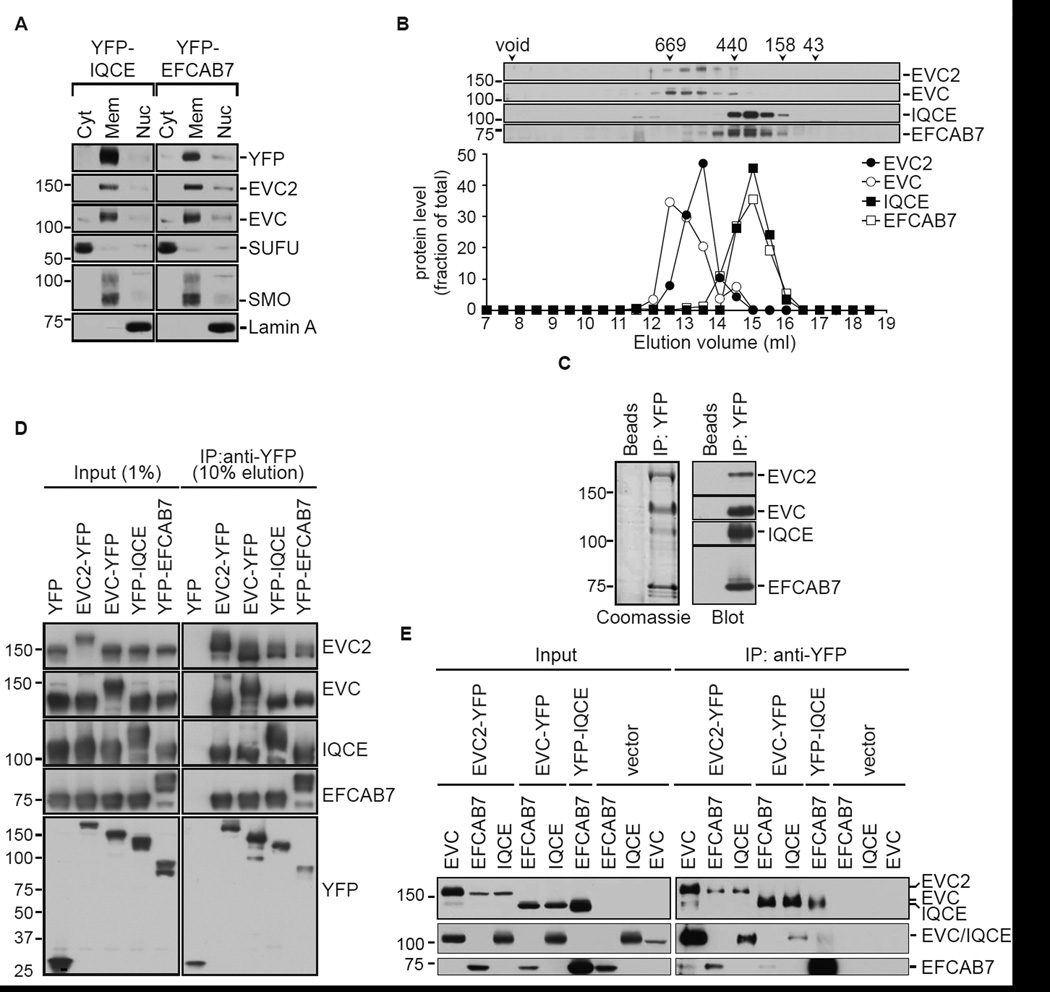
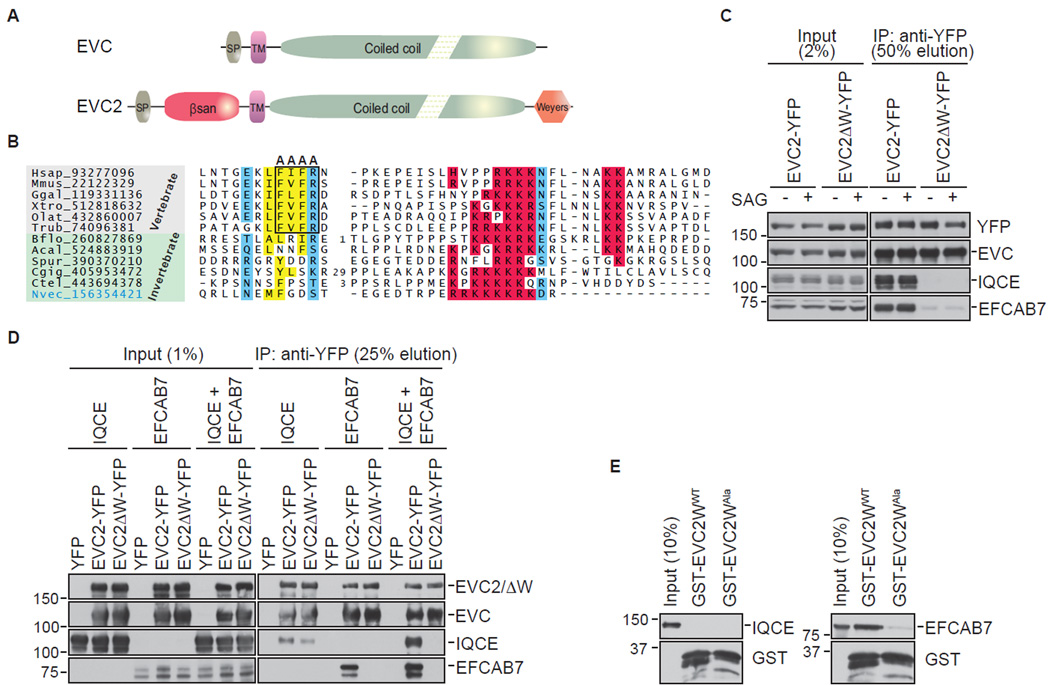
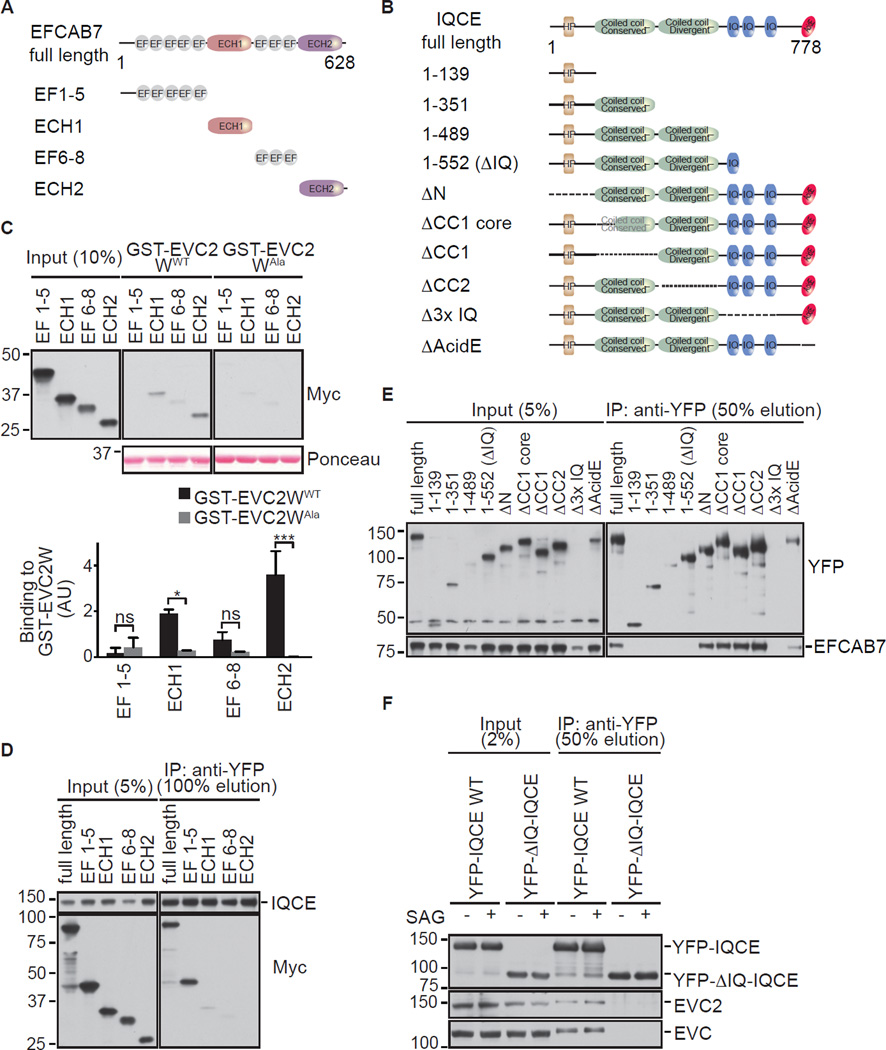
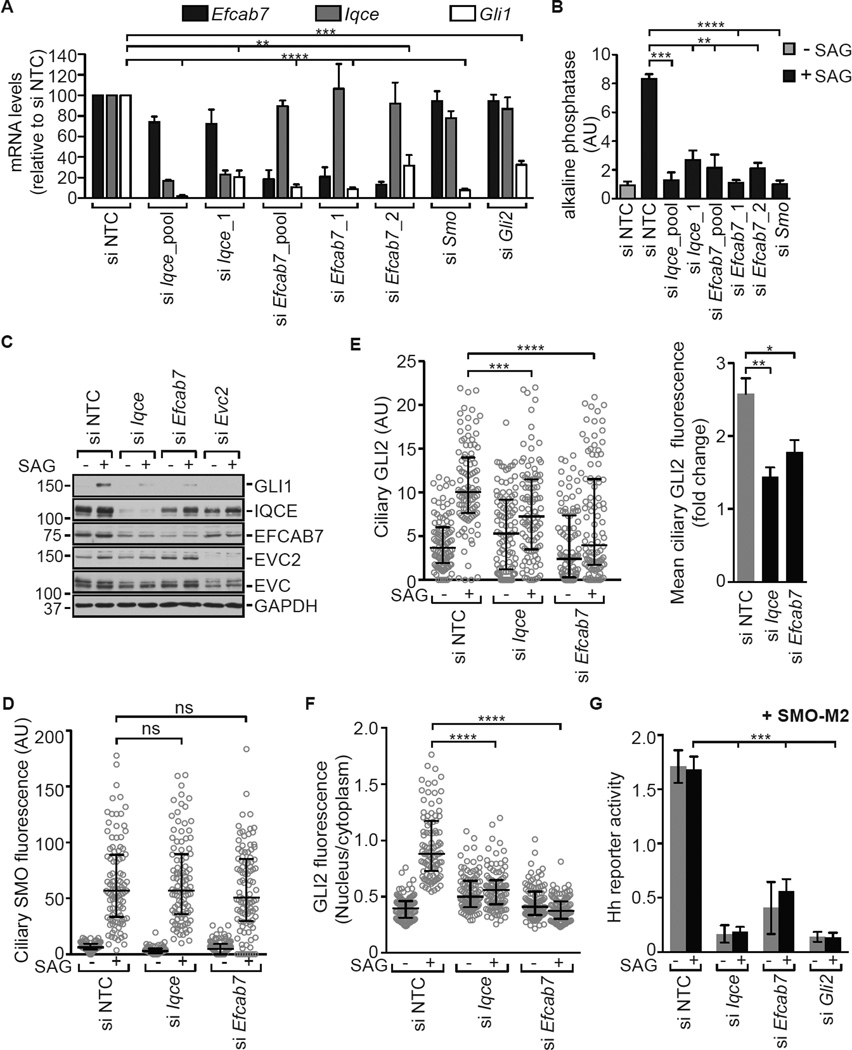
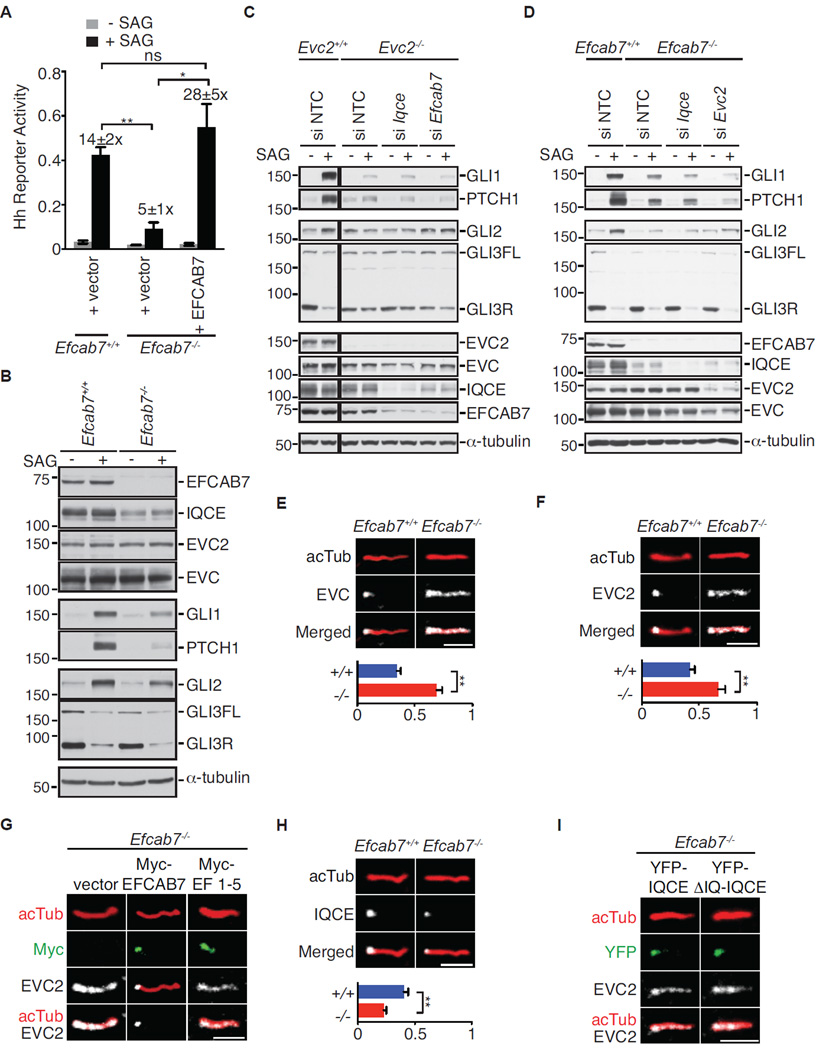
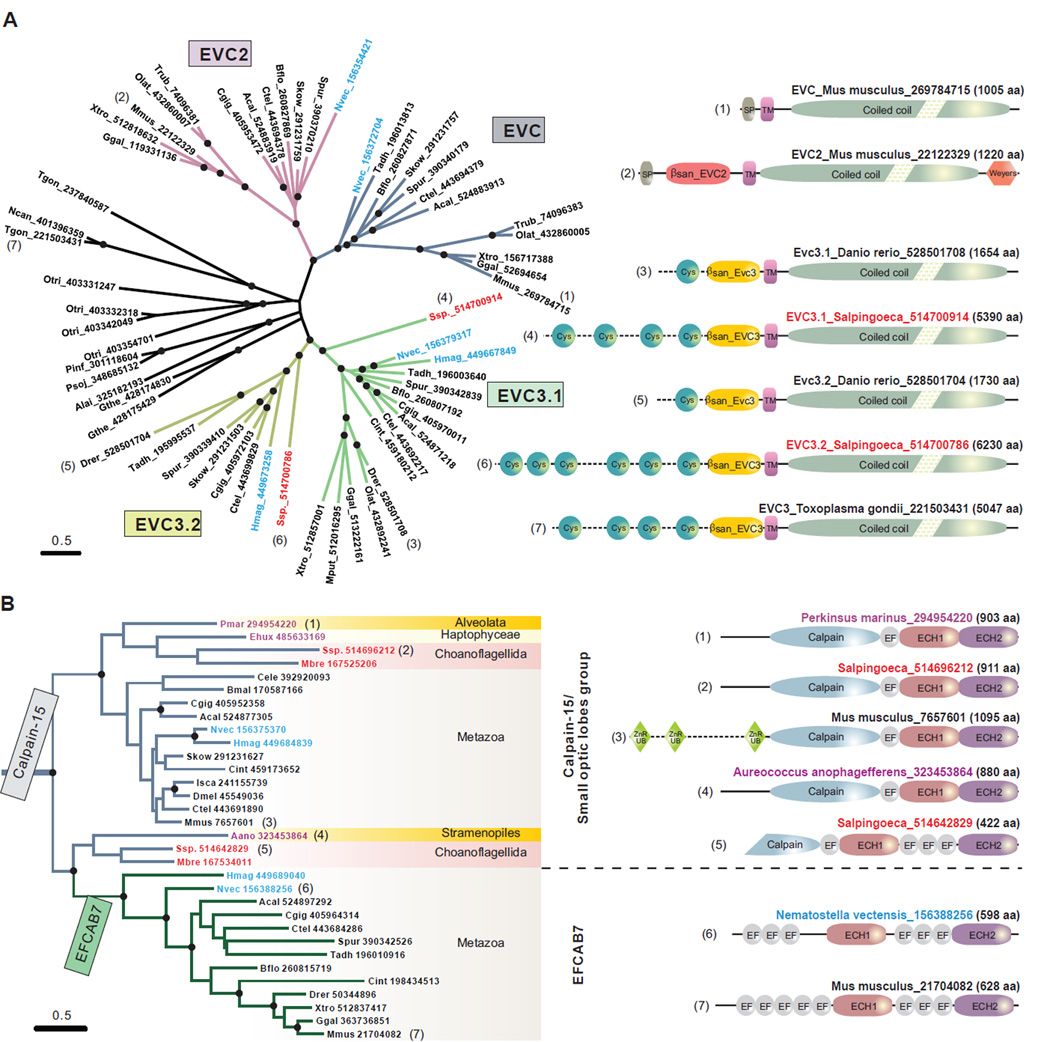
The Hedgehog (Hh) pathway depends on primary cilia in vertebrates, but the signaling machinery within cilia remains incompletely defined. We report the identification of a complex between two ciliary proteins, EFCAB7 and IQCE, which positively regulates the Hh pathway. The EFCAB7-IQCE module anchors the EVC-EVC2 complex in a signaling microdomain at the base of cilia. EVC and EVC2 genes are mutated in Ellis van Creveld and Weyers syndromes, characterized by impaired Hh signaling in skeletal, cardiac, and orofacial tissues. EFCAB7 binds to a C-terminal disordered region in EVC2 that is deleted in Weyers patients. EFCAB7 depletion mimics the Weyers cellular phenotype-the mislocalization of EVC-EVC2 within cilia and impaired activation of the transcription factor GLI2. Evolutionary analysis suggests that emergence of these complexes might have been important for adaptation of an ancient organelle, the cilium, for an animal-specific signaling network.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Smoothened transduces Hedgehog signal by forming a complex with Evc/Evc2.Cell Res. 2012 Nov;22(11):1593-604. doi: 10.1038/cr.2012.134. Epub 2012 Sep 18. Cell Res. 2012. PMID: 22986504 Free PMC article.
-
The ciliary Evc/Evc2 complex interacts with Smo and controls Hedgehog pathway activity in chondrocytes by regulating Sufu/Gli3 dissociation and Gli3 trafficking in primary cilia.Hum Mol Genet. 2013 Jan 1;22(1):124-39. doi: 10.1093/hmg/dds409. Epub 2012 Oct 1. Hum Mol Genet. 2013. PMID: 23026747
-
EVC-EVC2 complex stability and ciliary targeting are regulated by modification with ubiquitin and SUMO.Front Cell Dev Biol. 2023 Jul 27;11:1190258. doi: 10.3389/fcell.2023.1190258. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37576597 Free PMC article.
-
Ellis-van Creveld syndrome and Weyers acrodental dysostosis are caused by cilia-mediated diminished response to hedgehog ligands.Am J Med Genet C Semin Med Genet. 2009 Nov 15;151C(4):341-51. doi: 10.1002/ajmg.c.30226. Am J Med Genet C Semin Med Genet. 2009. PMID: 19876929 Review.
-
Role of primary cilia and Hedgehog signaling in craniofacial features of Ellis-van Creveld syndrome.Am J Med Genet C Semin Med Genet. 2022 Mar;190(1):36-46. doi: 10.1002/ajmg.c.31969. Epub 2022 Apr 8. Am J Med Genet C Semin Med Genet. 2022. PMID: 35393766 Review.
Cited by
-
G-protein-coupled receptors, Hedgehog signaling and primary cilia.Semin Cell Dev Biol. 2014 Sep;33:63-72. doi: 10.1016/j.semcdb.2014.05.002. Epub 2014 May 17. Semin Cell Dev Biol. 2014. PMID: 24845016 Free PMC article. Review.
-
Identification of a Novel IQCE Large Deletion through Copy Number Variant Analysis from Whole-Exome Sequencing Data of a Patient with Postaxial Polydactyly Type A7.Mol Syndromol. 2023 Jun;14(3):225-230. doi: 10.1159/000527777. Epub 2023 Jan 13. Mol Syndromol. 2023. PMID: 37323200 Free PMC article.
-
Novel mutations in EVC cause aberrant splicing in Ellis-van Creveld syndrome.Mol Genet Genomics. 2016 Apr;291(2):863-72. doi: 10.1007/s00438-015-1151-2. Epub 2015 Nov 30. Mol Genet Genomics. 2016. PMID: 26621368
-
Bacterial calpains and the evolution of the calpain (C2) family of peptidases.Biol Direct. 2015 Nov 2;10:66. doi: 10.1186/s13062-015-0095-0. Biol Direct. 2015. PMID: 26527411 Free PMC article.
-
Hedgehog signaling mechanism and role in cancer.Semin Cancer Biol. 2022 Oct;85:107-122. doi: 10.1016/j.semcancer.2021.04.003. Epub 2021 Apr 6. Semin Cancer Biol. 2022. PMID: 33836254 Free PMC article. Review.
References
-
- Caparros-Martin JA, Valencia M, Reytor E, Pacheco M, Fernandez M, Perez-Aytes A, Gean E, Lapunzina P, Peters H, Goodship JA, et al. The ciliary Evc/Evc2 complex interacts with Smo and controls Hedgehog pathway activity in chondrocytes by regulating Sufu/Gli3 dissociation and Gli3 trafficking in primary cilia. Hum Mol Genet. 2013;22:124–139. - PubMed
-
- Caspary T, Larkins CE, Anderson KV. The graded response to Sonic Hedgehog depends on cilia architecture. Dev Cell. 2007;12:767–778. - PubMed
-
- Corbit KC, Aanstad P, Singla V, Norman AR, Stainier DY, Reiter JF. Vertebrate Smoothened functions at the primary cilium. Nature. 2005;437:1018–1021. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
