

Functions of crystallins in and out of lens: roles in elongated and post-mitotic cells
- PMID: 24582830
- PMCID: PMC4104235
- DOI: 10.1016/j.pbiomolbio.2014.02.006
Functions of crystallins in and out of lens: roles in elongated and post-mitotic cells
Abstract
The vertebrate lens evolved to collect light and focus it onto the retina. In development, the lens grows through massive elongation of epithelial cells possibly recapitulating the evolutionary origins of the lens. The refractive index of the lens is largely dependent on high concentrations of soluble proteins called crystallins. All vertebrate lenses share a common set of crystallins from two superfamilies (although other lineage specific crystallins exist). The α-crystallins are small heat shock proteins while the β- and γ-crystallins belong to a superfamily that contains structural proteins of uncertain function. The crystallins are expressed at very high levels in lens but are also found at lower levels in other cells, particularly in retina and brain. All these proteins have plausible connections to maintenance of cytoplasmic order and chaperoning of the complex molecular machines involved in the architecture and function of cells, particularly elongated and post-mitotic cells. They may represent a suite of proteins that help maintain homeostasis in such cells that are at risk from stress or from the accumulated insults of aging.
Keywords: Chaperone; Crystallins; Epithelial cell; Lens; Retina; Stress.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
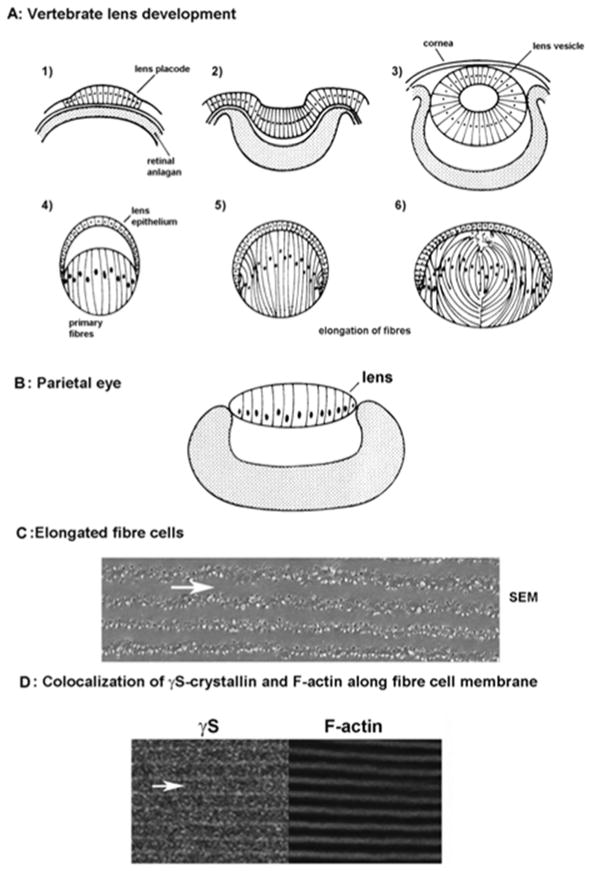
Epithelial cells overlying the retinal anlagen elongate to form the lens placode
Elongation continues as the optic vesicle invaginates.
Formation of the lens vesicle and cornea
Within the lens vesicle, primary fiber cells elongate
Fibres fill the lens
New layers of secondary fibres form throughout life through differentiation and elongation of equatorial epithelial cells.

References
-
- Acunzo J, Katsogiannou M, Rocchi P. Small heat shock proteins HSP27 (HspB1), alphaB-crystallin (HspB5) and HSP22 (HspB8) as regulators of cell death. Int J Biochem Cell Biol. 2012;44:1622–31. - PubMed
-
- Anderson DH, Stern WH, Fisher SK, Erickson PA, Borgula GA. The onset of pigment epithelial proliferation after retinal detachment. Invest Ophthalmol Vis Sci. 1981;21:10–6. - PubMed
-
- Andley UP. Crystallins in the eye: Function and pathology. Prog Retin Eye Res. 2007;26:78–98. - PubMed
-
- Arrigo AP. Human small heat shock proteins: protein interactomes of homo- and hetero-oligomeric complexes: an update. FEBS Lett. 2013;587:1959–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
