

Cooperative interaction of Etv2 and Gata2 regulates the development of endothelial and hematopoietic lineages
- PMID: 24583263
- PMCID: PMC4099474
- DOI: 10.1016/j.ydbio.2014.02.018
Cooperative interaction of Etv2 and Gata2 regulates the development of endothelial and hematopoietic lineages
Abstract
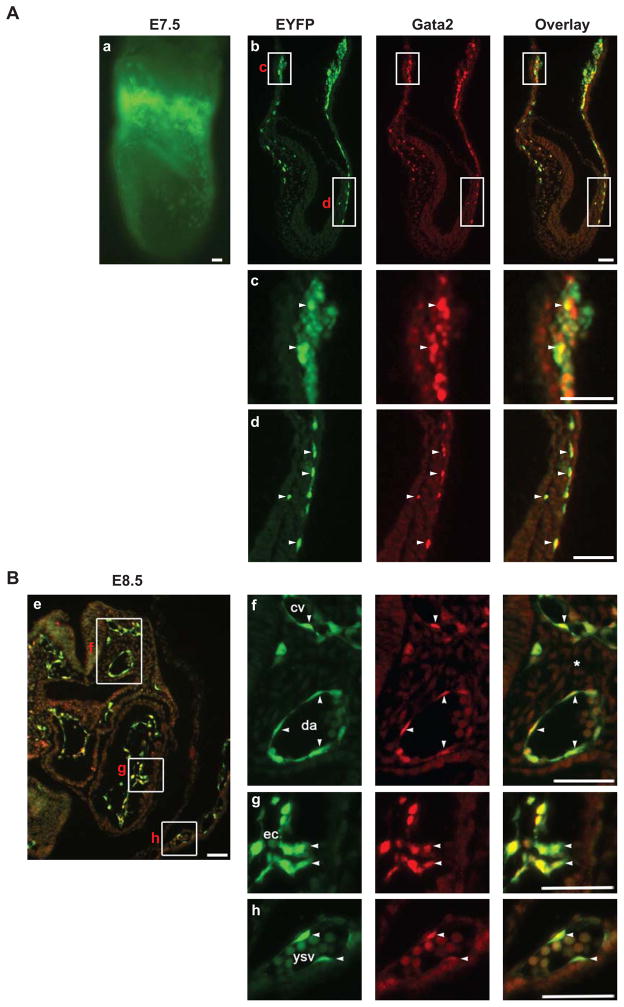
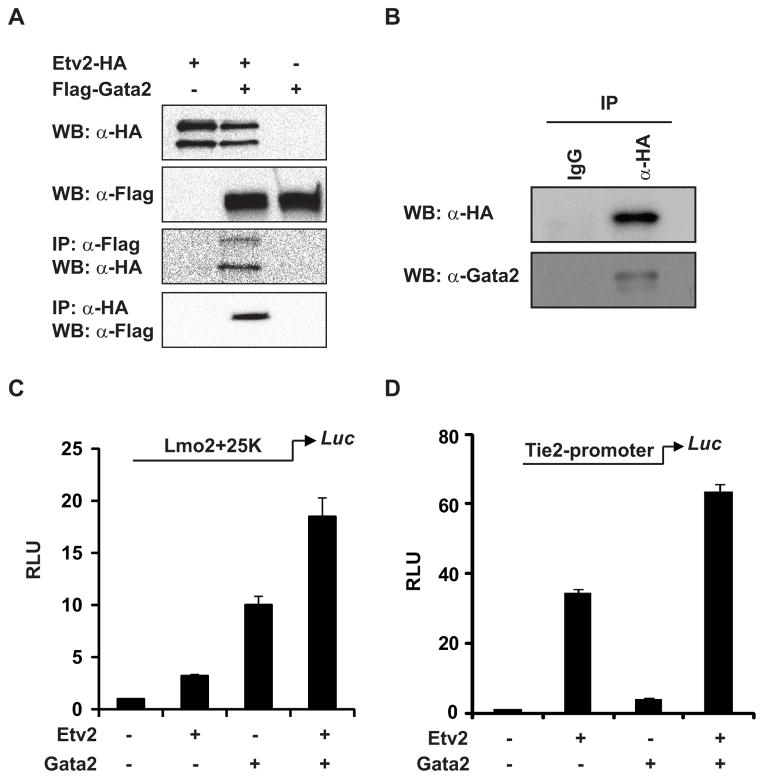
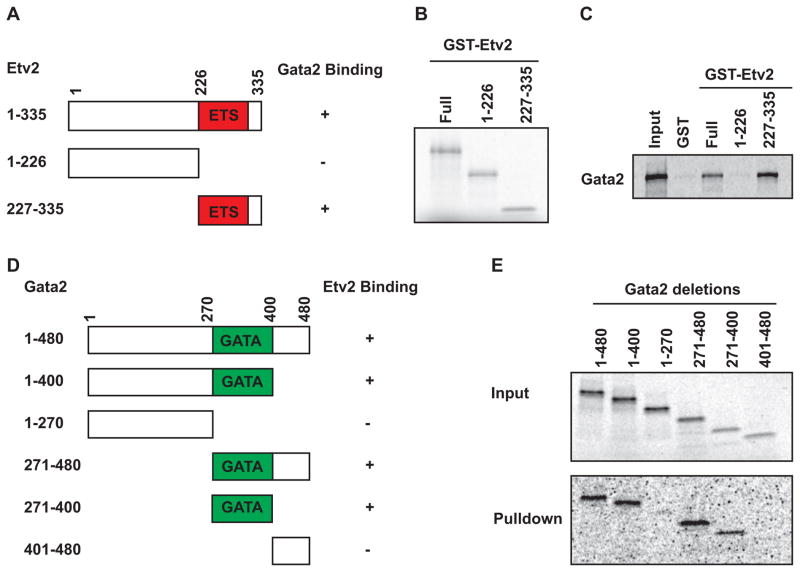
Regulatory mechanisms that govern lineage specification of the mesodermal progenitors to become endothelial and hematopoietic cells remain an area of intense interest. Both Ets and Gata factors have been shown to have important roles in the transcriptional regulation in endothelial and hematopoietic cells. We previously reported Etv2 as an essential regulator of vasculogenesis and hematopoiesis. In the present study, we demonstrate that Gata2 is co-expressed and interacts with Etv2 in the endothelial and hematopoietic cells in the early stages of embryogenesis. Our studies reveal that Etv2 interacts with Gata2 in vitro and in vivo. The protein-protein interaction between Etv2 and Gata2 is mediated by the Ets and Gata domains. Using the embryoid body differentiation system, we demonstrate that co-expression of Gata2 augments the activity of Etv2 in promoting endothelial and hematopoietic lineage differentiation. We also identify Spi1 as a common downstream target gene of Etv2 and Gata2. We provide evidence that Etv2 and Gata2 bind to the Spi1 promoter in vitro and in vivo. In summary, we propose that Gata2 functions as a cofactor of Etv2 in the transcriptional regulation of mesodermal progenitors during embryogenesis.
Keywords: Endothelial cell; Etv2; Gata2; Hematopoietic cell.
Copyright © 2014. Published by Elsevier Inc.
Figures


References
-
- Bories JC, Willerford DM, Grevin D, Davidson L, Camus A, Martin P, Stehelin D, Alt FW. Increased T-cell apoptosis and terminal B-cell differentiation induced by inactivation of the Ets-1 proto-oncogene. Nature. 1995;377:635–638. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
