

Themes and variations in riboswitch structure and function
- PMID: 24583553
- PMCID: PMC4643838
- DOI: 10.1016/j.bbagrm.2014.02.012
Themes and variations in riboswitch structure and function
Abstract
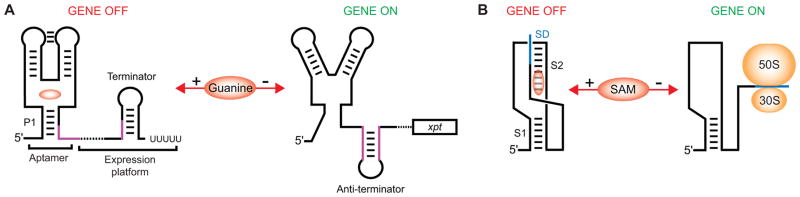
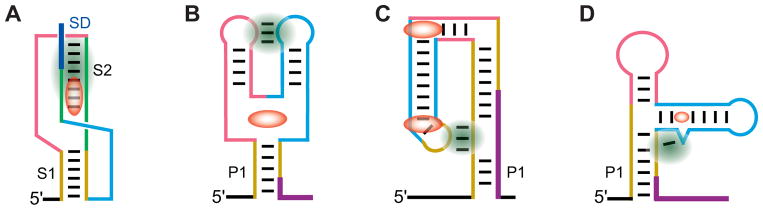
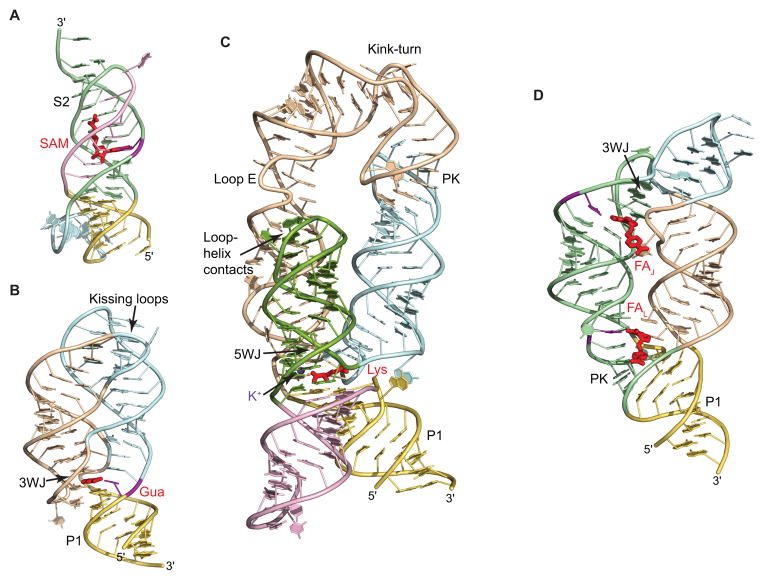
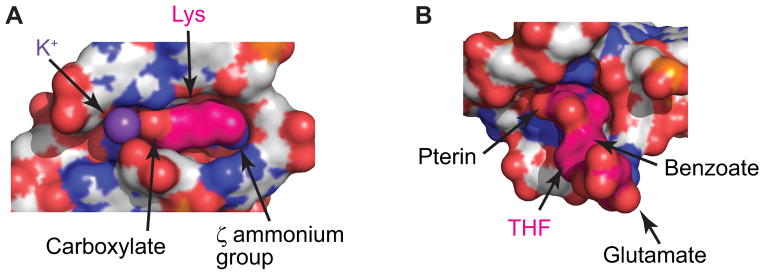
The complexity of gene expression control by non-coding RNA has been highlighted by the recent progress in the field of riboswitches. Discovered a decade ago, riboswitches represent a diverse group of non-coding mRNA regions that possess a unique ability to directly sense cellular metabolites and modulate gene expression through formation of alternative metabolite-free and metabolite-bound conformations. Such protein-free metabolite sensing domains utilize sophisticated three-dimensional folding of RNA molecules to discriminate between a cognate ligand from related compounds so that only the right ligand would trigger a genetic response. Given the variety of riboswitch ligands ranging from small cations to large coenzymes, riboswitches adopt a great diversity of structures. Although many riboswitches share structural principles to build metabolite-competent folds, form precise ligand-binding pockets, and communicate a ligand-binding event to downstream regulatory regions, virtually all riboswitch classes possess unique features for ligand recognition, even those tuned to recognize the same metabolites. Here we present an overview of the biochemical and structural research on riboswitches with a major focus on common principles and individual characteristics adopted by these regulatory RNA elements during evolution to specifically target small molecules and exert genetic responses. This article is part of a Special Issue entitled: Riboswitches.
Keywords: Gene expression; Metabolite; Non-coding RNA; RNA structure; Transcription; X-ray crystallography.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
-
- Kim SH, Suddath FL, Quigley GJ, McPherson A, Sussman JL, Wang AH, Seeman NC, Rich A. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science. 1974;185:435–440. - PubMed
-
- Cech TR, Zaug AJ, Grabowski PJ. In vitro splicing of the ribosomal RNA precursor of Tetrahymena: involvement of a guanosine nucleotide in the excision of the intervening sequence. Cell. 1981;27:487–496. - PubMed
-
- Ellington AD, Szostak JW. In vitro selection of RNA molecules that bind specific ligands. Nature. 1990;346:818–822. - PubMed
-
- Tuerk C, Gold L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science. 1990;249:505–510. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
