

Review
doi: 10.1152/physiol.00050.2013.
AMPK: regulating energy balance at the cellular and whole body levels
Affiliations
- PMID: 24583766
- PMCID: PMC3949207
- DOI: 10.1152/physiol.00050.2013
Item in Clipboard
Review
AMPK: regulating energy balance at the cellular and whole body levels
Physiology (Bethesda).
2014 Mar.
Abstract
AMP-activated protein kinase appears to have evolved in single-celled eukaryotes as an adenine nucleotide sensor that maintains energy homeostasis at the cellular level. However, during evolution of more complex multicellular organisms, the system has adapted to interact with hormones so that it also plays a key role in balancing energy intake and expenditure at the whole body level.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures
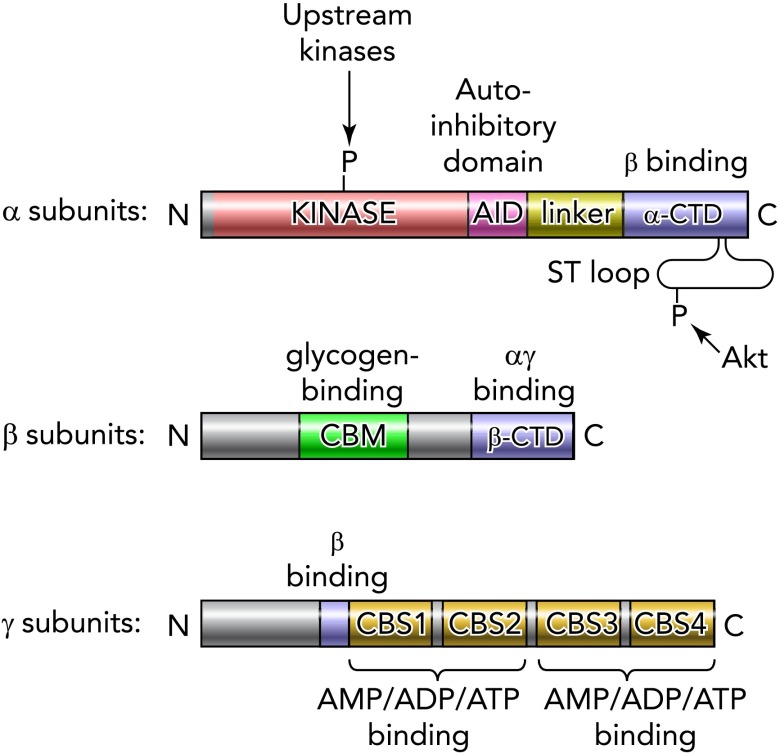
Domain structure of eukaryotic AMPK orthologs The catalytic α-subunits contain conventional serine/threonine kinase domains containing the threonine residue (Thr172 in rat α2) phosphorylated by upstream kinases. The kinase domains are followed (at least in vertebrates) by small domains with a negative effect on kinase activity (auto-inhibitory domains), which are joined to the COOH-terminal domains (α-CTD) by a less well conserved linker. In vertebrates, there is also a flexible serine-/threonine-rich loop (ST loop) within the α-CTD that is phosphorylated by Akt. The β-subunits contain two conserved regions, a carbohydrate-binding module (CBM) that causes the mammalian complex to bind to glycogen particles, and a COOH-terminal domain (β-CTD) that provides the bridge between the α- and γ-subunits. The γ-subunits contain variable NH2-terminal regions followed by a short sequence involved in binding to the β-subunit, then four tandem repeats of a cystathionine-β-synthase (CBS) motif. These act in pairs to form the binding sites for adenine nucleotides; in mammalian AMPK, there is one site between CBS1 and CBS2 and two between CBS3 and CBS4.
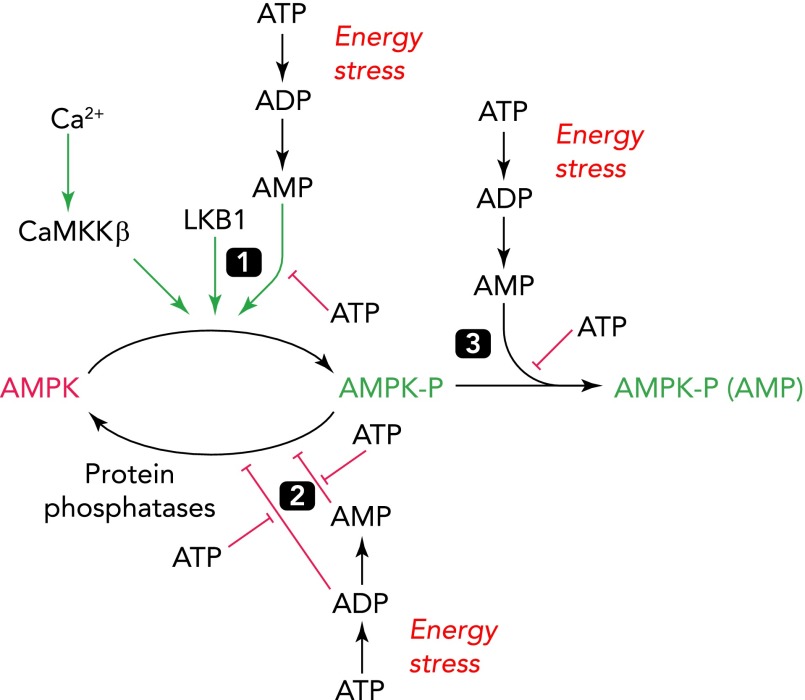
Regulation of mammalian AMPK by adenine nucleotides and Ca2+ Cellular energy stress leads to net conversion of ATP to ADP, some of which is converted to AMP by the adenylate kinase reaction (2ADP ↔ ATP + AMP), which maintains AMP at low levels in unstressed cells. AMP then activates AMPK by three mechanisms, all of which are due to binding to one or more sites on the AMPK-γ subunit: 1) promoting Thr172 phosphorylation by LKB1; 2) inhibiting Thr172 dephosphorylation by protein phosphatases; 3) allosteric activation. Only mechanism 2 is mimicked by ADP, and then only at higher concentrations than AMP (20). AMPK can also be activated by increases in cytosolic Ca2+, which triggers direct phosphorylation of Thr172 by CaMKKβ.
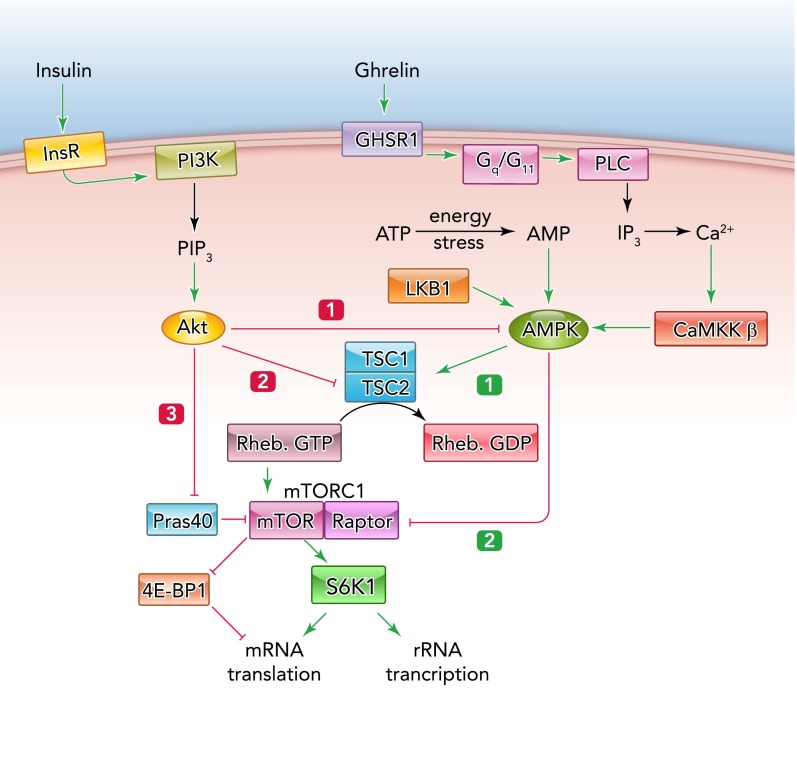
Regulation of mTORC1 by the insulin/Akt and AMPK signaling pathways The insulin receptor activates phosphoinositide 3-kinase (PI3K), causing production of phosphatidylinositol 3,4,5-trisphosphate (PIP3), the second messenger that switches on the protein kinase Akt. Akt then has at least three effects: 1) it phosphorylates AMPK, antagonizing its activation by LKB1; 2) it phosphorylates TSC2 at sites that oppose its function as a Rheb-GAP, thus activating mTORC1; 3) it phosphorylates PRAS40, relieving its inhibitory effects on mTORC1. mTORC1 (a complex containing mTOR, Raptor, and other components) then phosphorylates S6K1 and 4E-BP1, promoting cell growth by enhancing translation of specific mRNAs, as well as rRNA synthesis. By contrast, AMPK, which is activated by energy stress or by hormones that increase cytosolic Ca2+ such as ghrelin, has two effects: 1) it phosphorylates TSC2 at distinct sites, enhancing its Rheb-GAP activity; 2) it phosphorylates Raptor. These effects act to inhibit mTORC1, protein synthesis, and cell growth.
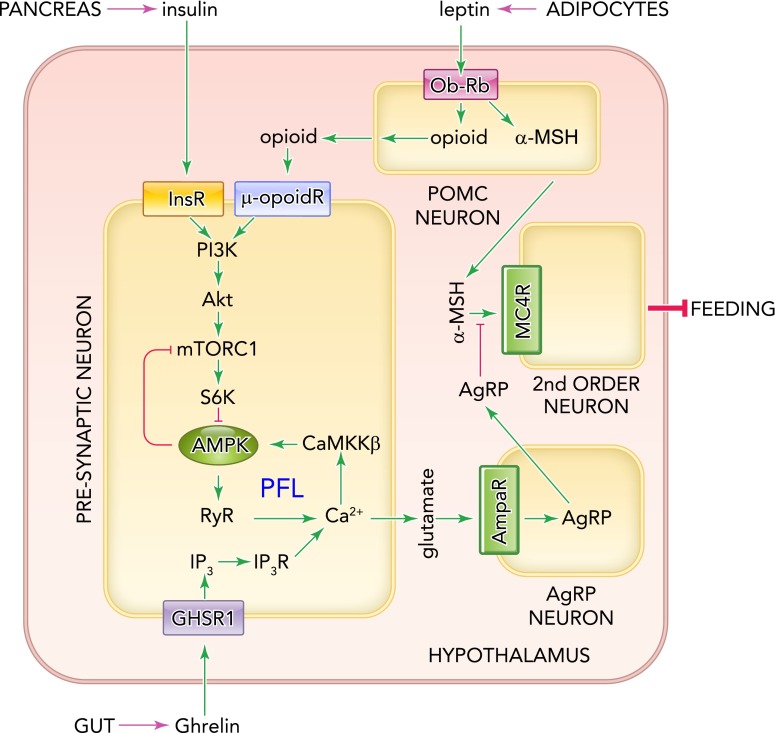
Model for role of AMPK in controlling feeding behavior in mammals, based on Sternson (81) and Kahn (15) According to this combined model, the key role of AMPK is in presynaptic neurons immediately upstream of NPY/AgRP neurons in the hypothalamus. Ghrelin causes release of inositol 3,4,5-triphosphate (IP3) in these cells via the G-protein-coupled receptor GHSR1, leading to release of Ca2+ via IP3, thus activating CaMKKβ and hence AMPK (one effect of which is to inhibit mTORC1 via the mechanisms shown in FIGURE 3). The released Ca2+ also triggers release of glutamate that activates the downstream NPY/AgRP neurons, initiating feeding via release of AgRP onto second-order neurons. AMPK is also proposed to activate ryanodine receptors (RyR) that release Ca2+, setting up a positive feedback loop (PFL) that allows continuous stimulation of the NPY/AgRP neuron even if stimulation by ghrelin ceases. This loop is interrupted and feeding stops when POMC neurons release an opioid onto the presynaptic neuron, inhibiting AMPK via the PI3K-Akt-mTORC1-S6K1 pathway. Insulin also depresses feeding by activating the same pathway in the presynaptic neuron.
Similar articles
-
AMP-activated protein kinase: maintaining energy homeostasis at the cellular and whole-body levels.Annu Rev Nutr. 2014;34:31-55. doi: 10.1146/annurev-nutr-071812-161148. Epub 2014 May 15. Annu Rev Nutr. 2014. PMID: 24850385 Free PMC article. Review.
-
AMPK: a metabolic gauge regulating whole-body energy homeostasis.Trends Mol Med. 2008 Dec;14(12):539-49. doi: 10.1016/j.molmed.2008.09.007. Epub 2008 Nov 1. Trends Mol Med. 2008. PMID: 18977694 Review.
-
AMPK: a nutrient and energy sensor that maintains energy homeostasis.Nat Rev Mol Cell Biol. 2012 Mar 22;13(4):251-62. doi: 10.1038/nrm3311. Nat Rev Mol Cell Biol. 2012. PMID: 22436748 Free PMC article. Review.
-
Hypothalamic AMPK as a Regulator of Energy Homeostasis.Neural Plast. 2016;2016:2754078. doi: 10.1155/2016/2754078. Epub 2016 Jul 28. Neural Plast. 2016. PMID: 27547453 Free PMC article. Review.
-
Hypothalamic AMPK as a Mediator of Hormonal Regulation of Energy Balance.Int J Mol Sci. 2018 Nov 11;19(11):3552. doi: 10.3390/ijms19113552. Int J Mol Sci. 2018. PMID: 30423881 Free PMC article. Review.
Cited by
-
Mice deficient in the mitochondrial branched-chain aminotransferase (BCATm) respond with delayed tumour growth to a challenge with EL-4 lymphoma.Br J Cancer. 2018 Oct;119(8):1009-1017. doi: 10.1038/s41416-018-0283-7. Epub 2018 Oct 15. Br J Cancer. 2018. PMID: 30318512 Free PMC article.
-
Transcriptomic and Metabolomic Data Reveal the Key Metabolic Pathways Affecting Streltzoviella insularis (Staudinger) (Lepidoptera: Cossidae) Larvae During Overwintering.Front Physiol. 2021 Jun 18;12:655059. doi: 10.3389/fphys.2021.655059. eCollection 2021. Front Physiol. 2021. PMID: 34220530 Free PMC article.
-
A review of the basics of mitochondrial bioenergetics, metabolism, and related signaling pathways in cancer cells: Therapeutic targeting of tumor mitochondria with lipophilic cationic compounds.Redox Biol. 2018 Apr;14:316-327. doi: 10.1016/j.redox.2017.09.020. Epub 2017 Sep 29. Redox Biol. 2018. PMID: 29017115 Free PMC article. Review.
-
The role of declining adaptive homeostasis in ageing.J Physiol. 2017 Dec 15;595(24):7275-7309. doi: 10.1113/JP275072. Epub 2017 Nov 21. J Physiol. 2017. PMID: 29028112 Free PMC article. Review.
-
Effect of different γ-subunit isoforms on the regulation of AMPK.Biochem J. 2017 May 9;474(10):1741-1754. doi: 10.1042/BCJ20170046. Biochem J. 2017. PMID: 28302767 Free PMC article.
References
-
- Anderson KA, Ribar TJ, Lin F, Noeldner PK, Green MF, Muehlbauer MJ, Witters LA, Kemp BE, Means AR. Hypothalamic CaMKK2 contributes to the regulation of energy balance. Cell Metab 7: 377–388, 2008 - PubMed
-
- Andersson U, Filipsson K, Abbott CR, Woods A, Smith K, Bloom SR, Carling D, Small CJ. AMP-activated protein kinase plays a role in the control of food intake. J Biol Chem 279: 12005–12008, 2004 - PubMed
-
- Baena-Gonzalez E, Rolland F, Thevelein JM, Sheen J. A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942, 2007 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources