

Association of the breast cancer antiestrogen resistance protein 1 (BCAR1) and BCAR3 scaffolding proteins in cell signaling and antiestrogen resistance
- PMID: 24584939
- PMCID: PMC4036165
- DOI: 10.1074/jbc.M113.541839
Association of the breast cancer antiestrogen resistance protein 1 (BCAR1) and BCAR3 scaffolding proteins in cell signaling and antiestrogen resistance
Abstract
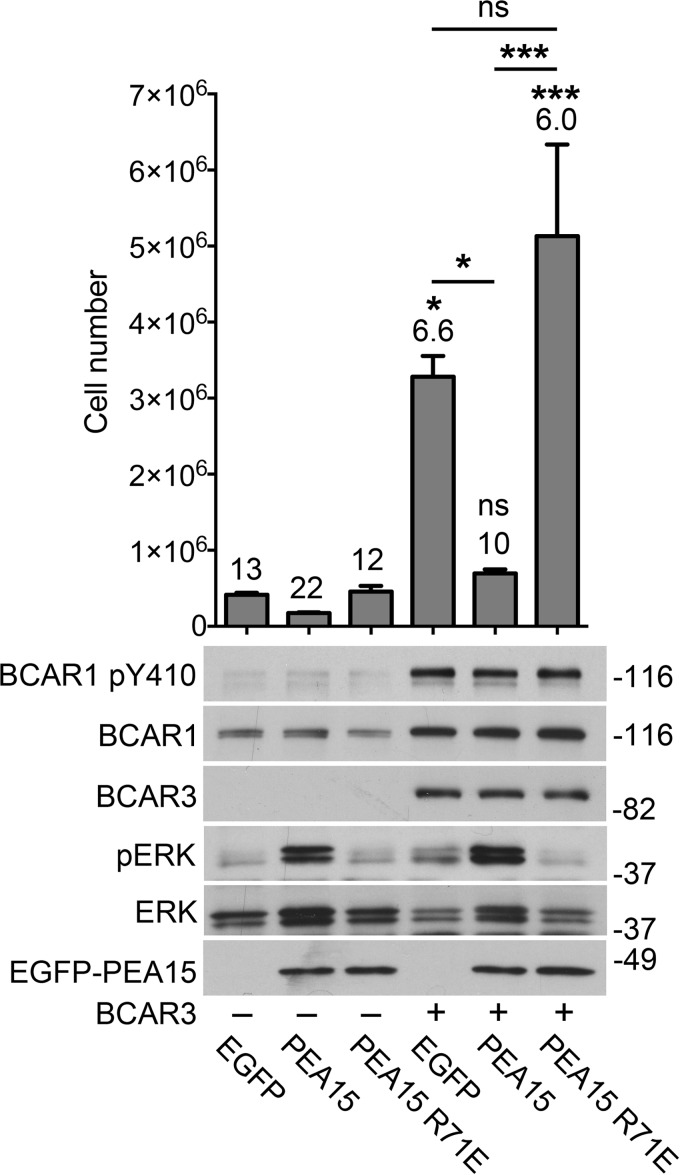
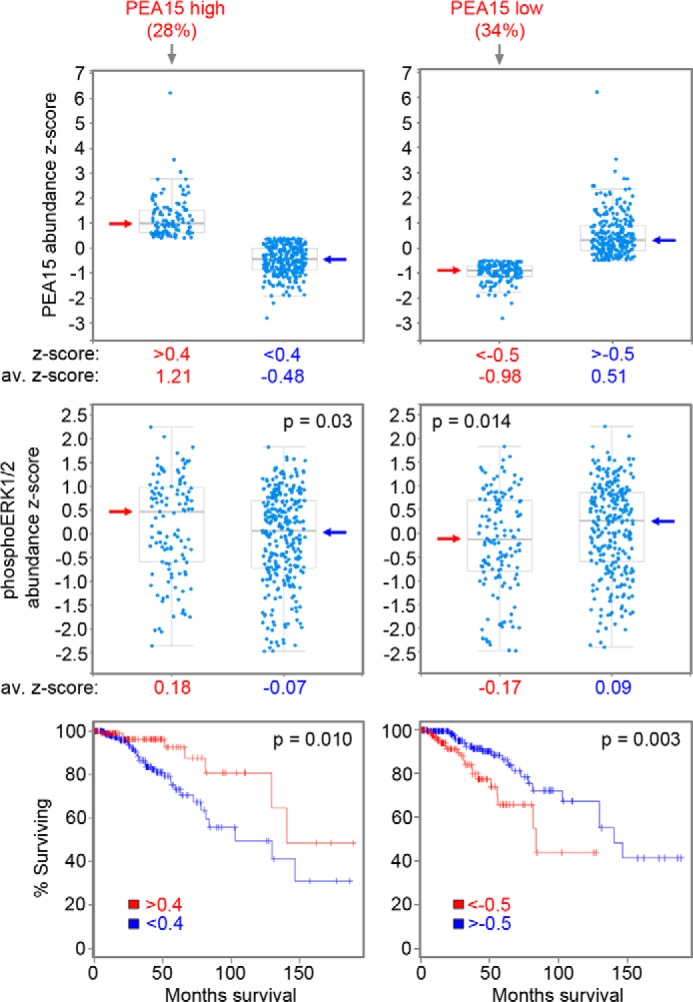
Most breast cancers are estrogen receptor-positive and treated with antiestrogens, but aberrant signaling networks can induce drug resistance. One of these networks involves the scaffolding protein BCAR1/p130CAS, which regulates cell growth and migration/invasion. A less investigated scaffolding protein that also confers antiestrogen resistance is the SH2 domain-containing protein BCAR3. BCAR1 and BCAR3 bind tightly to each other through their C-terminal domains, thus potentially connecting their associated signaling networks. However, recent studies using BCAR1 and BCAR3 interaction mutants concluded that association between the two proteins is not critical for many of their interrelated activities regulating breast cancer malignancy. We report that these previously used BCAR mutations fail to cause adequate loss-of-function of the complex. By using structure-based BCAR1 and BCAR3 mutants that lack the ability to interact, we show that BCAR3-induced antiestrogen resistance in MCF7 breast cancer cells critically depends on its ability to bind BCAR1. Interaction with BCAR3 increases the levels of phosphorylated BCAR1, ultimately potentiating BCAR1-dependent antiestrogen resistance. Furthermore, antiestrogen resistance in cells overexpressing BCAR1/BCAR3 correlates with increased ERK1/2 activity. Inhibiting ERK1/2 through overexpression of the regulatory protein PEA15 negates the resistance, revealing a key role for ERK1/2 in BCAR1/BCAR3-induced antiestrogen resistance. Reverse-phase protein array data show that PEA15 levels in invasive breast cancers correlate with patient survival, suggesting that PEA15 can override ERK1/2 activation by BCAR1/BCAR3 and other upstream regulators. We further uncovered that the BCAR3-related NSP3 can also promote antiestrogen resistance. Thus, strategies to disrupt BCAR1-BCAR3/NSP3 complexes and associated signaling networks could ultimately lead to new breast cancer therapies.
Keywords: Anticancer Drug; Breast Cancer; Cancer; Drug Resistance; ERK1/2; MAP Kinases (MAPKs); Mesenchymal Phenotype; NSP3; PEA15; Protein-Protein Interactions.
Figures
Similar articles
-
AND-34/BCAR3 differs from other NSP homologs in induction of anti-estrogen resistance, cyclin D1 promoter activation and altered breast cancer cell morphology.J Cell Physiol. 2007 Sep;212(3):655-65. doi: 10.1002/jcp.21059. J Cell Physiol. 2007. PMID: 17427198 Free PMC article.
-
Protein pathway activation mapping reveals molecular networks associated with antiestrogen resistance in breast cancer cell lines.Int J Cancer. 2012 Nov 1;131(9):1998-2007. doi: 10.1002/ijc.27489. Epub 2012 Mar 20. Int J Cancer. 2012. PMID: 22328489
-
AND-34/BCAR3, a GDP exchange factor whose overexpression confers antiestrogen resistance, activates Rac, PAK1, and the cyclin D1 promoter.Cancer Res. 2003 Oct 15;63(20):6802-8. Cancer Res. 2003. PMID: 14583477
-
Antiestrogen resistance in breast cancer and the role of estrogen receptor signaling.Oncogene. 2003 Oct 20;22(47):7316-39. doi: 10.1038/sj.onc.1206937. Oncogene. 2003. PMID: 14576841 Review.
-
Inhibition of erbB receptor (HER) tyrosine kinases as a strategy to abrogate antiestrogen resistance in human breast cancer.Clin Cancer Res. 2001 Dec;7(12 Suppl):4436s-4442s; discussion 4411s-4412s. Clin Cancer Res. 2001. PMID: 11916237 Review.
Cited by
-
Candidate biomarkers for treatment benefit from sunitinib in patients with advanced renal cell carcinoma using mass spectrometry-based (phospho)proteomics.Clin Proteomics. 2023 Nov 8;20(1):49. doi: 10.1186/s12014-023-09437-6. Clin Proteomics. 2023. PMID: 37940875 Free PMC article.
-
A novel spontaneous mutation of BCAR3 results in extrusion cataracts in CF#1 mouse strain.Mamm Genome. 2016 Oct;27(9-10):451-9. doi: 10.1007/s00335-016-9653-8. Epub 2016 Jun 30. Mamm Genome. 2016. PMID: 27364350
-
Mechanisms of tRNA-derived fragments and tRNA halves in cancer treatment resistance.Biomark Res. 2020 Oct 15;8:52. doi: 10.1186/s40364-020-00233-0. eCollection 2020. Biomark Res. 2020. PMID: 33072328 Free PMC article. Review.
-
Altered regulation of PDK4 expression promotes antiestrogen resistance in human breast cancer cells.Springerplus. 2015 Nov 10;4:689. doi: 10.1186/s40064-015-1444-2. eCollection 2015. Springerplus. 2015. PMID: 26576332 Free PMC article.
-
BCAR3 promotes head and neck cancer growth and is associated with poor prognosis.Cell Death Discov. 2021 Oct 27;7(1):316. doi: 10.1038/s41420-021-00714-7. Cell Death Discov. 2021. PMID: 34707118 Free PMC article.
References
-
- Musgrove E. A., Sutherland R. L. (2009) Biological determinants of endocrine resistance in breast cancer. Nat. Rev. Cancer 9, 631–643 - PubMed
-
- Brinkman A., van der Flier S., Kok E. M., Dorssers L. C. (2000) BCAR1, a human homologue of the adapter protein p130Cas, and antiestrogen resistance in breast cancer cells. J. Natl. Cancer Inst. 92, 112–120 - PubMed
-
- van der Flier S., Chan C. M., Brinkman A., Smid M., Johnston S. R., Dorssers L. C., Dowsett M. (2000) BCAR1/p130Cas expression in untreated and acquired tamoxifen-resistant human breast carcinomas. Int. J. Cancer 89, 465–468 - PubMed
-
- van der Flier S., Brinkman A., Look M. P., Kok E. M., Meijer-van Gelder M. E., Klijn J. G., Dorssers L. C., Foekens J. A. (2000) Bcar1/p130Cas protein and primary breast cancer. Prognosis and response to tamoxifen treatment. J. Natl. Cancer Inst. 92, 120–127 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
