

Phage lysis: three steps, three choices, one outcome
- PMID: 24585055
- PMCID: PMC4012431
- DOI: 10.1007/s12275-014-4087-z
Phage lysis: three steps, three choices, one outcome
Abstract
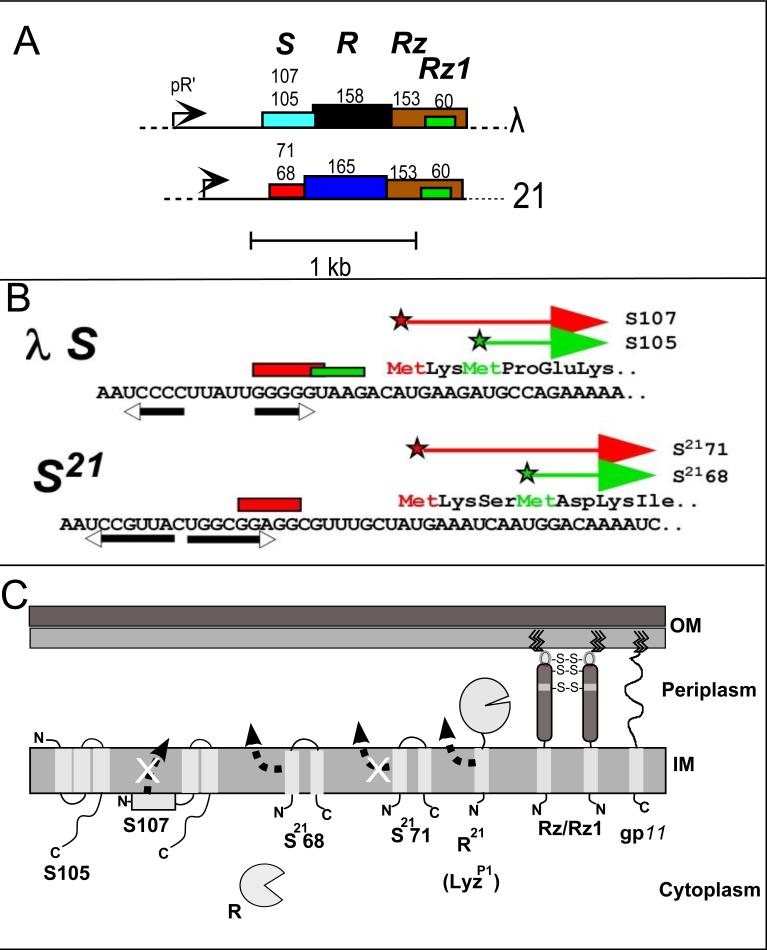
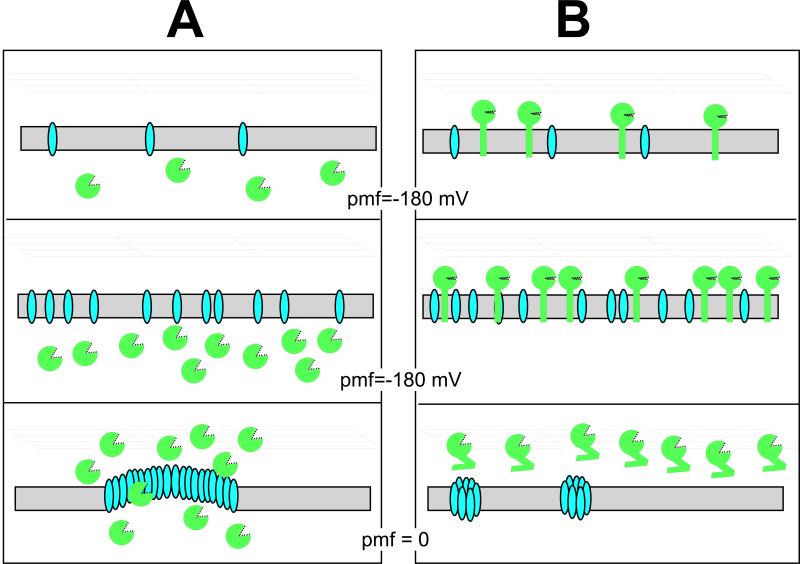
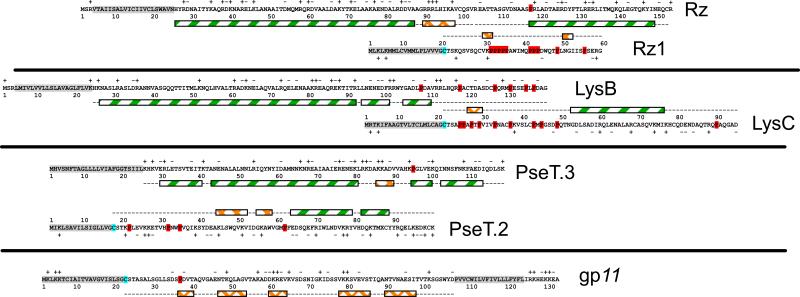
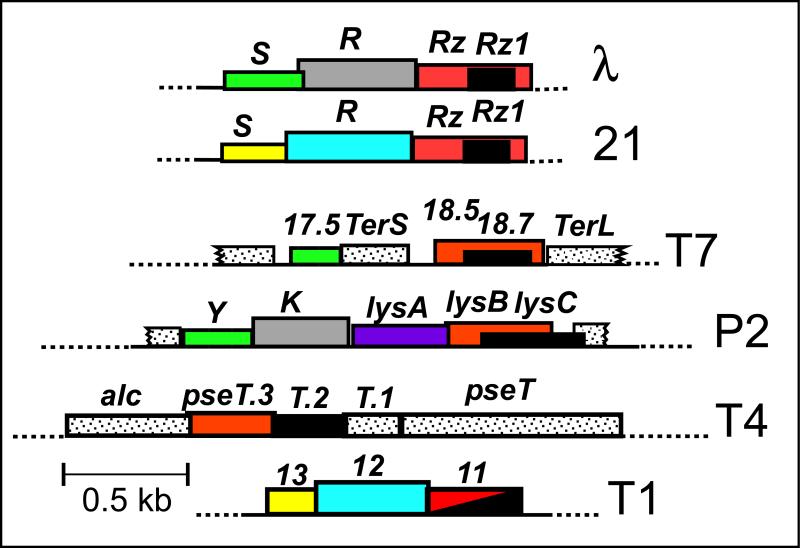
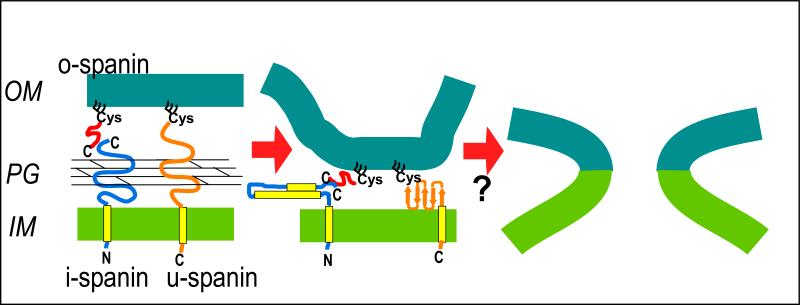
The lysis of bacterial hosts by double-strand DNA bacteriophages, once thought to reflect merely the accumulation of sufficient lysozyme activity during the infection cycle, has been revealed to recently been revealed to be a carefully regulated and temporally scheduled process. For phages of Gramnegative hosts, there are three steps, corresponding to subversion of each of the three layers of the cell envelope: inner membrane, peptidoglycan, and outer membrane. The pathway is controlled at the level of the cytoplasmic membrane. In canonical lysis, a phage encoded protein, the holin, accumulates harmlessly in the cytoplasmic membrane until triggering at an allele-specific time to form micron-scale holes. This allows the soluble endolysin to escape from the cytoplasm to degrade the peptidoglycan. Recently a parallel pathway has been elucidated in which a different type of holin, the pinholin, which, instead of triggering to form large holes, triggers to form small, heptameric channels that serve to depolarize the membrane. Pinholins are associated with SAR endolysins, which accumulate in the periplasm as inactive, membrane-tethered enzymes. Pinholin triggering collapses the proton motive force, allowing the SAR endolysins to refold to an active form and attack the peptidoglycan. Surprisingly, a third step, the disruption of the outer membrane is also required. This is usually achieved by a spanin complex, consisting of a small outer membrane lipoprotein and an integral cytoplasmic membrane protein, designated as o-spanin and i-spanin, respectively. Without spanin function, lysis is blocked and progeny virions are trapped in dead spherical cells, suggesting that the outer membrane has considerable tensile strength. In addition to two-component spanins, there are some single-component spanins, or u-spanins, that have an N-terminal outer-membrane lipoprotein signal and a C-terminal transmembrane domain. A possible mechanism for spanin function to disrupt the outer membrane is to catalyze fusion of the inner and outer membranes.
Figures

References
-
- Adhya S, Sen A, Mitra S. In: The role of gene S In: The Bacteriophage Lambda. Hershey AD, editor. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: 1971. pp. 743–746.
-
- Atkins JF, Steitz JA, Anderson CW, Model P. Binding of mammalian ribosomes to MS2 phage RNA reveals an overlapping gene encoding a lysis function. Cell. 1979;18:247–256. - PubMed
-
- Barenboim M, Chang CY, dib Hajj F, Young R. Characterization of the dual start motif of a class II holin gene. Mol.Microbiol. 1999;32:715–727. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
