

tRNA signatures reveal a polyphyletic origin of SAR11 strains among alphaproteobacteria
- PMID: 24586126
- PMCID: PMC3937112
- DOI: 10.1371/journal.pcbi.1003454
tRNA signatures reveal a polyphyletic origin of SAR11 strains among alphaproteobacteria
Abstract
Molecular phylogenetics and phylogenomics are subject to noise from horizontal gene transfer (HGT) and bias from convergence in macromolecular compositions. Extensive variation in size, structure and base composition of alphaproteobacterial genomes has complicated their phylogenomics, sparking controversy over the origins and closest relatives of the SAR11 strains. SAR11 are highly abundant, cosmopolitan aquatic Alphaproteobacteria with streamlined, A+T-biased genomes. A dominant view holds that SAR11 are monophyletic and related to both Rickettsiales and the ancestor of mitochondria. Other studies dispute this, finding evidence of a polyphyletic origin of SAR11 with most strains distantly related to Rickettsiales. Although careful evolutionary modeling can reduce bias and noise in phylogenomic inference, entirely different approaches may be useful to extract robust phylogenetic signals from genomes. Here we develop simple phyloclassifiers from bioinformatically derived tRNA Class-Informative Features (CIFs), features predicted to target tRNAs for specific interactions within the tRNA interaction network. Our tRNA CIF-based model robustly and accurately classifies alphaproteobacterial genomes into one of seven undisputed monophyletic orders or families, despite great variability in tRNA gene complement sizes and base compositions. Our model robustly rejects monophyly of SAR11, classifying all but one strain as Rhizobiales with strong statistical support. Yet remarkably, conventional phylogenetic analysis of tRNAs classifies all SAR11 strains identically as Rickettsiales. We attribute this discrepancy to convergence of SAR11 and Rickettsiales tRNA base compositions. Thus, tRNA CIFs appear more robust to compositional convergence than tRNA sequences generally. Our results suggest that tRNA-CIF-based phyloclassification is robust to HGT of components of the tRNA interaction network, such as aminoacyl-tRNA synthetases. We explain why tRNAs are especially advantageous for prediction of traits governing macromolecular interactions from genomic data, and why such traits may be advantageous in the search for robust signals to address difficult problems in classification and phylogeny.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
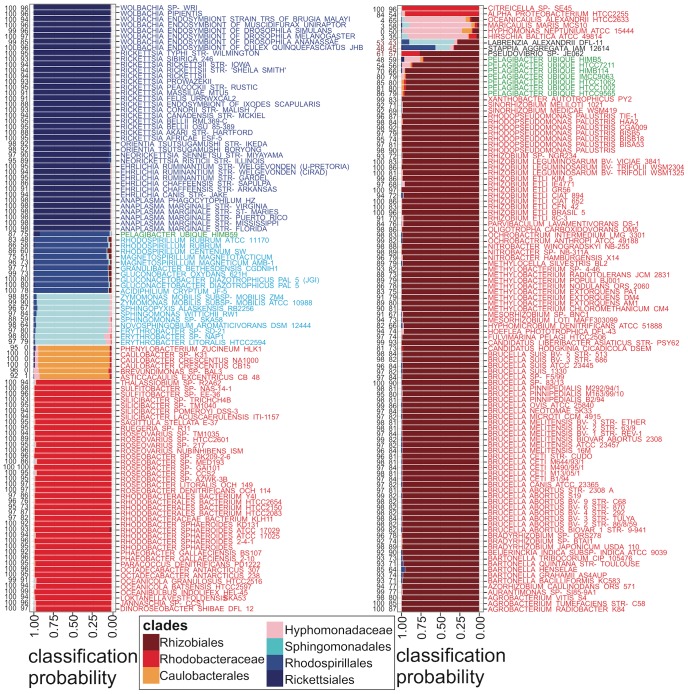
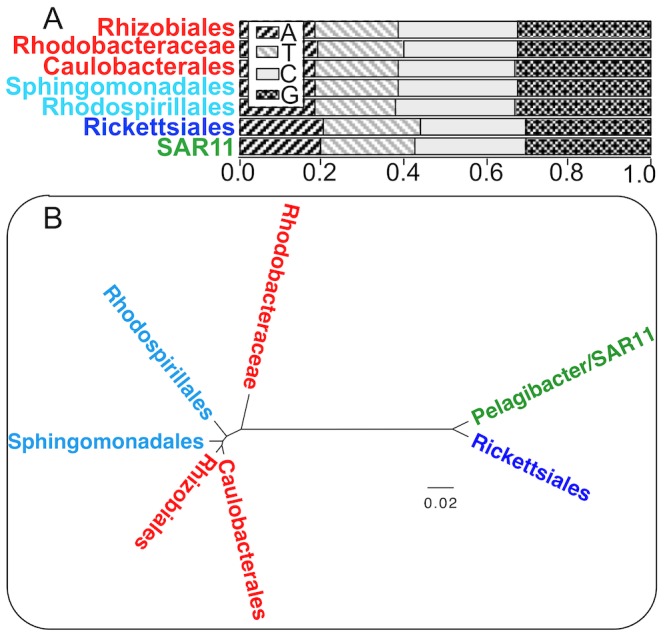
 bits and model retraining (100 replicates). All support values correspond to most probable clade as shown except for Stappia and Labrenzia for which they correspond to Rhizobiales. Complete source code and data to produce this figure, including the full WEKA model for classification of other alphaproteobacterial genomes and code to produce such models from scratch, is provided in Dataset S4.
bits and model retraining (100 replicates). All support values correspond to most probable clade as shown except for Stappia and Labrenzia for which they correspond to Rhizobiales. Complete source code and data to produce this figure, including the full WEKA model for classification of other alphaproteobacterial genomes and code to produce such models from scratch, is provided in Dataset S4.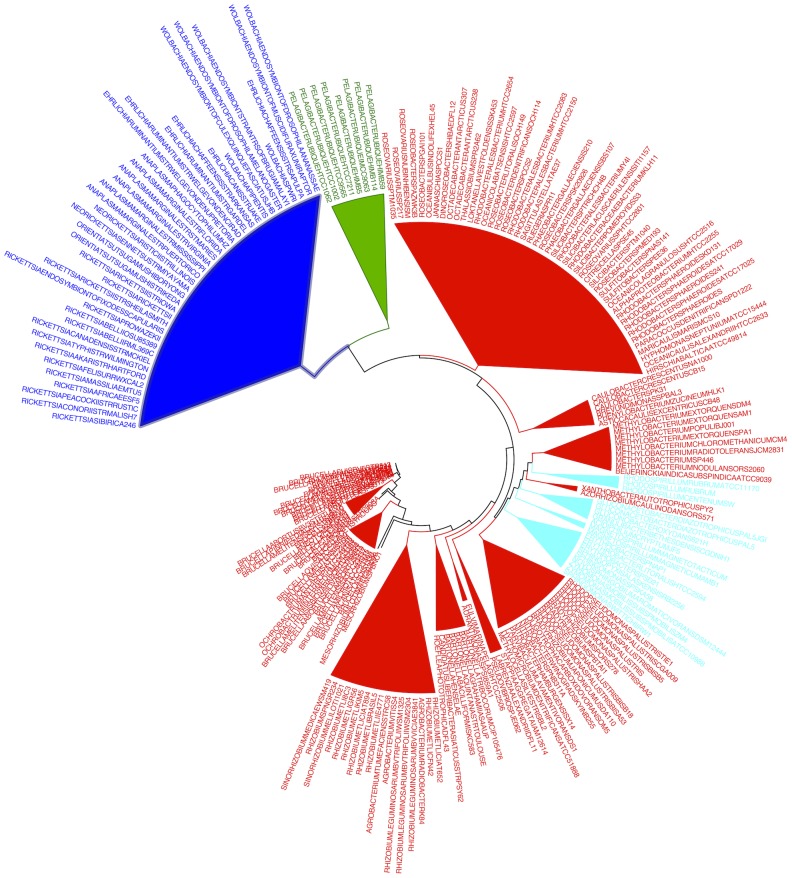
References
-
- Gribaldo S, Philippe H (2002) Ancient phylogenetic relationships. Theor Popul Biol 61: 391–408. - PubMed
-
- Gogarten JP, Doolittle WF, Lawrence JG (2002) Prokaryotic evolution in light of gene transfer. Mol Biol Evol 19: 2226–2238. - PubMed
-
- Morris RM, Rappé MS, Connon SA, Vergin KL, Siebold WA, et al. (2002) SAR 11 clade dominates ocean surface bacterioplankton communities. Nature 420: 806–810. - PubMed
-
- Giovannoni SJ (2005) Genome streamlining in a cosmopolitan oceanic bacterium. Science 309: 1242–1245. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
