

A unified model of heading and path perception in primate MSTd
- PMID: 24586130
- PMCID: PMC3930491
- DOI: 10.1371/journal.pcbi.1003476
A unified model of heading and path perception in primate MSTd
Abstract
Self-motion, steering, and obstacle avoidance during navigation in the real world require humans to travel along curved paths. Many perceptual models have been proposed that focus on heading, which specifies the direction of travel along straight paths, but not on path curvature, which humans accurately perceive and is critical to everyday locomotion. In primates, including humans, dorsal medial superior temporal area (MSTd) has been implicated in heading perception. However, the majority of MSTd neurons respond optimally to spiral patterns, rather than to the radial expansion patterns associated with heading. No existing theory of curved path perception explains the neural mechanisms by which humans accurately assess path and no functional role for spiral-tuned cells has yet been proposed. Here we present a computational model that demonstrates how the continuum of observed cells (radial to circular) in MSTd can simultaneously code curvature and heading across the neural population. Curvature is encoded through the spirality of the most active cell, and heading is encoded through the visuotopic location of the center of the most active cell's receptive field. Model curvature and heading errors fit those made by humans. Our model challenges the view that the function of MSTd is heading estimation, based on our analysis we claim that it is primarily concerned with trajectory estimation and the simultaneous representation of both curvature and heading. In our model, temporal dynamics afford time-history in the neural representation of optic flow, which may modulate its structure. This has far-reaching implications for the interpretation of studies that assume that optic flow is, and should be, represented as an instantaneous vector field. Our results suggest that spiral motion patterns that emerge in spatio-temporal optic flow are essential for guiding self-motion along complex trajectories, and that cells in MSTd are specifically tuned to extract complex trajectory estimation from flow.
Conflict of interest statement
Dr. Browning is now a paid employee of Scientific Systems Company Inc. (SSCI). Dr. Browning was employed by Boston University while the research for the present work was conducted.
Figures


 , the monkey views a radially expanding optic flow while moving CCW when the heading direction is straight ahead, which is the same as the optic flow viewed CW
, the monkey views a radially expanding optic flow while moving CCW when the heading direction is straight ahead, which is the same as the optic flow viewed CW  on the other side of the circle. Between
on the other side of the circle. Between  and
and  , the FoE drifts rightward until at
, the FoE drifts rightward until at  it is out of view. At
it is out of view. At  , the monkey experiences radial contraction.
, the monkey experiences radial contraction.



 outside the path. (c) On path condition. The observer maintained gaze on a target on the future path positioned
outside the path. (c) On path condition. The observer maintained gaze on a target on the future path positioned  from the initial heading. (d) Inside path condition. Observer gaze was maintained on a target positioned
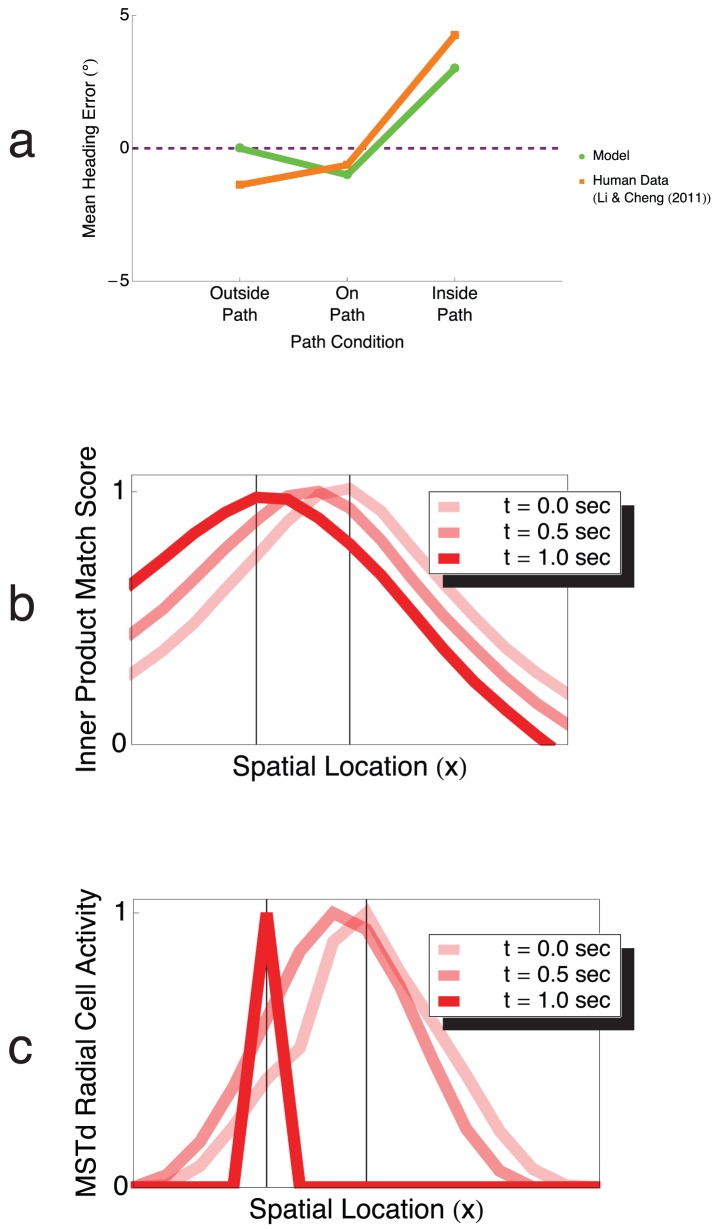
from the initial heading. (d) Inside path condition. Observer gaze was maintained on a target positioned  inside the path. (e) Gaze along heading condition. Observer gaze is always tangent to the circular path, which is most often the case during human locomotion. Human subjects in the experiments of Li and Cheng underestimated path curvature in the Z axis, outside path, and on path conditions, overestimated path curvature in the inside path condition, and yielded low error in their judgments in the gaze along heading condition.
inside the path. (e) Gaze along heading condition. Observer gaze is always tangent to the circular path, which is most often the case during human locomotion. Human subjects in the experiments of Li and Cheng underestimated path curvature in the Z axis, outside path, and on path conditions, overestimated path curvature in the inside path condition, and yielded low error in their judgments in the gaze along heading condition.
 path radius for the five gaze conditions. The location of each peak across the spiral continuum determines the model estimate of path curvature. For example, in the Z-axis condition (black), the MSTd activity peak occurs in the subpopulation sensitive to radial expansion (
path radius for the five gaze conditions. The location of each peak across the spiral continuum determines the model estimate of path curvature. For example, in the Z-axis condition (black), the MSTd activity peak occurs in the subpopulation sensitive to radial expansion ( ), and therefore the model indicates zero path curvature (straight path). (c) Model path errors (solid lines) compared to human data from Li and Cheng (replotted, dashed lines) in the five gaze conditions as a function of path curvature . Model path errors were in good agreement in all gaze conditions with those based on human judgments (
), and therefore the model indicates zero path curvature (straight path). (c) Model path errors (solid lines) compared to human data from Li and Cheng (replotted, dashed lines) in the five gaze conditions as a function of path curvature . Model path errors were in good agreement in all gaze conditions with those based on human judgments ( ), and path error decreased linearly (
), and path error decreased linearly ( ) with path curvature. Error bars correspond to standard error of the mean (SEM).
) with path curvature. Error bars correspond to standard error of the mean (SEM).


 (a),
(a),  (b),
(b),  (c) radius paths. Lesions were introduced in the model MSTd connectivity by zeroing out competitive interactions in spiral space, across spiral orientation, and across 2D space between neurons in MSTd (see Eq. 6). Lesions had a detrimental impact on model performance, and path curvature estimates no longer mapped onto human judgments. The three competitive interactions in model MSTd were necessary to obtain our results.
(c) radius paths. Lesions were introduced in the model MSTd connectivity by zeroing out competitive interactions in spiral space, across spiral orientation, and across 2D space between neurons in MSTd (see Eq. 6). Lesions had a detrimental impact on model performance, and path curvature estimates no longer mapped onto human judgments. The three competitive interactions in model MSTd were necessary to obtain our results.
 (a),
(a),  (b), and
(b), and  (c), respectively. Deviations in path error were modest, with mean errors falling within
(c), respectively. Deviations in path error were modest, with mean errors falling within  of those depicted in Figure 7. There were only small deviations in any condition when the scene contained at least
of those depicted in Figure 7. There were only small deviations in any condition when the scene contained at least  .
.
 , dark orange), model MSTd neurons most sensitive to spiral patterns were most active. This also occurred when for larger angular rotation rates (
, dark orange), model MSTd neurons most sensitive to spiral patterns were most active. This also occurred when for larger angular rotation rates ( ). When the angular rotation was set to a comparable rate to that used in the Z-axis condition (
). When the angular rotation was set to a comparable rate to that used in the Z-axis condition ( ), model MSTd neurons most sensitive to radial expansion elicited the maximal activation. (b) Simplified model mechanisms that explain why neurons that are sensitive to spirals produced the peak activity in spiral space in the simulation of the Froehler and Duffy experiment, but did not in the simulation of the Z-axis condition. Consider the first-order optic flow (
), model MSTd neurons most sensitive to radial expansion elicited the maximal activation. (b) Simplified model mechanisms that explain why neurons that are sensitive to spirals produced the peak activity in spiral space in the simulation of the Froehler and Duffy experiment, but did not in the simulation of the Z-axis condition. Consider the first-order optic flow ( and
and  ) at two times (
) at two times ( and
and  ) during the circular path traversal (top row). Template matching in the model is inversely weighted by distance to the FoE or CoM (second row). The third row shows the optimal templates inversely weighted by distance (
) during the circular path traversal (top row). Template matching in the model is inversely weighted by distance to the FoE or CoM (second row). The third row shows the optimal templates inversely weighted by distance ( and
and  ). Because model MSTd dynamically integrates afferent signals from model MT, activation due to the input at
). Because model MSTd dynamically integrates afferent signals from model MT, activation due to the input at  influences the activation due to the input at
influences the activation due to the input at  . Temporal accumulation in the model can be approximated by considering
. Temporal accumulation in the model can be approximated by considering  , which temporally blends the two weighted fields. This yields a spiral field (bottom row), and explains why model MSTd neurons sensitive to spiral patterns are most active when the angular rotation rate about the circular path is sufficiently large.
, which temporally blends the two weighted fields. This yields a spiral field (bottom row), and explains why model MSTd neurons sensitive to spiral patterns are most active when the angular rotation rate about the circular path is sufficiently large.Similar articles
-
Estimating curvilinear self-motion from optic flow with a biologically inspired neural system.Bioinspir Biomim. 2022 Jun 9;17(4). doi: 10.1088/1748-3190/ac709b. Bioinspir Biomim. 2022. PMID: 35580573
-
3D Visual Response Properties of MSTd Emerge from an Efficient, Sparse Population Code.J Neurosci. 2016 Aug 10;36(32):8399-415. doi: 10.1523/JNEUROSCI.0396-16.2016. J Neurosci. 2016. PMID: 27511012 Free PMC article.
-
Distributed encoding of curvilinear self-motion across spiral optic flow patterns.Sci Rep. 2022 Aug 4;12(1):13393. doi: 10.1038/s41598-022-16371-4. Sci Rep. 2022. PMID: 35927277 Free PMC article.
-
Visual and vestibular cue integration for heading perception in extrastriate visual cortex.J Physiol. 2011 Feb 15;589(Pt 4):825-33. doi: 10.1113/jphysiol.2010.194720. Epub 2010 Aug 2. J Physiol. 2011. PMID: 20679353 Free PMC article. Review.
-
Computational mechanisms for optic flow analysis in primate cortex.Int Rev Neurobiol. 2000;44:235-68. doi: 10.1016/s0074-7742(08)60745-x. Int Rev Neurobiol. 2000. PMID: 10605649 Review. No abstract available.
Cited by
-
Hierarchical structure is employed by humans during visual motion perception.Proc Natl Acad Sci U S A. 2020 Sep 29;117(39):24581-24589. doi: 10.1073/pnas.2008961117. Epub 2020 Sep 16. Proc Natl Acad Sci U S A. 2020. PMID: 32938799 Free PMC article.
-
A Neural Model of MST and MT Explains Perceived Object Motion during Self-Motion.J Neurosci. 2016 Aug 3;36(31):8093-102. doi: 10.1523/JNEUROSCI.4593-15.2016. J Neurosci. 2016. PMID: 27488630 Free PMC article.
-
Cortical Motion Perception Emerges from Dimensionality Reduction with Evolved Spike-Timing-Dependent Plasticity Rules.J Neurosci. 2022 Jul 27;42(30):5882-5898. doi: 10.1523/JNEUROSCI.0384-22.2022. Epub 2022 Jun 22. J Neurosci. 2022. PMID: 35732492 Free PMC article.
-
Self-motion perception in the elderly.Front Hum Neurosci. 2014 Sep 15;8:681. doi: 10.3389/fnhum.2014.00681. eCollection 2014. Front Hum Neurosci. 2014. PMID: 25309379 Free PMC article.
-
A neural mechanism for optic flow parsing in macaque visual cortex.Curr Biol. 2024 Nov 4;34(21):4983-4997.e9. doi: 10.1016/j.cub.2024.09.030. Epub 2024 Oct 9. Curr Biol. 2024. PMID: 39389059
References
-
- Gibson JJ (1979) The Ecological Approach To Visual Perception. Psychology Press : 1–174.
-
- Warren WH (1998) The State of Flow. In: Watanabe T, editor, High-level motion processing, Cambridge: MIT Press. pp. 315–358.
-
- Warren WHW, Morris MWM, Kalish MM (1988) Perception of translational heading from optical flow. Journal of experimental psychology Human perception and performance 14: 646–660. - PubMed
-
- van den Berg AVA (1992) Robustness of perception of heading from optic flow. Vision research 32: 1285–1296. - PubMed
-
- van den Berg AV (1996) Judgements of heading. Vision research 36: 2337–2350. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
