

Structure of the membrane anchor of pestivirus glycoprotein E(rns), a long tilted amphipathic helix
- PMID: 24586172
- PMCID: PMC3937272
- DOI: 10.1371/journal.ppat.1003973
Structure of the membrane anchor of pestivirus glycoprotein E(rns), a long tilted amphipathic helix
Abstract
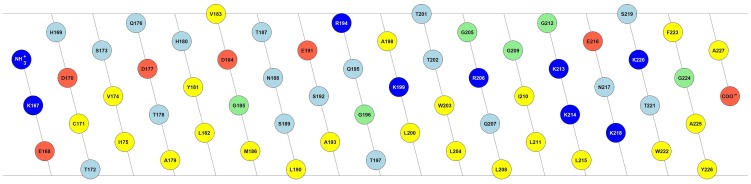
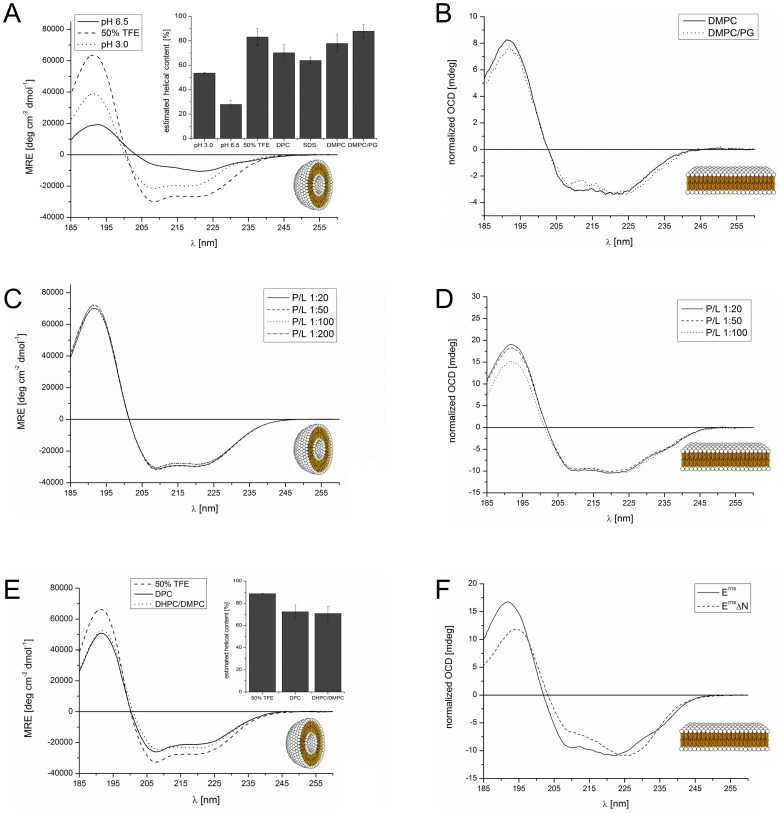
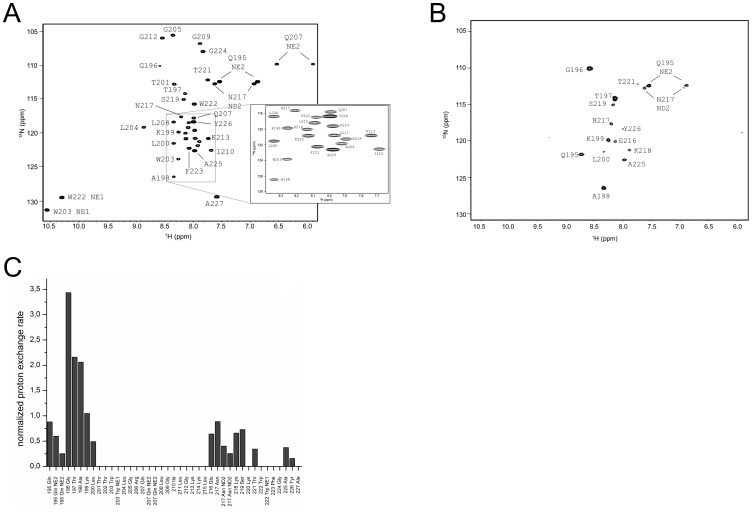
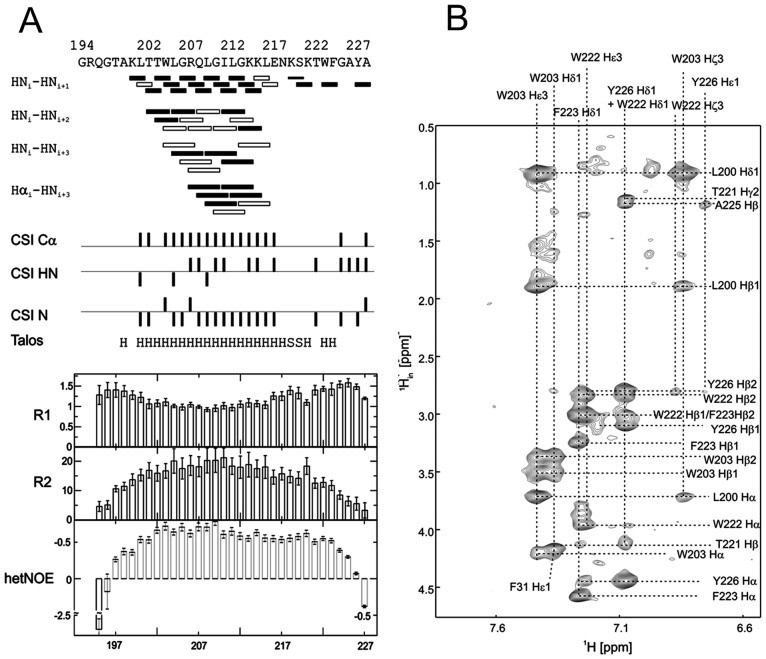
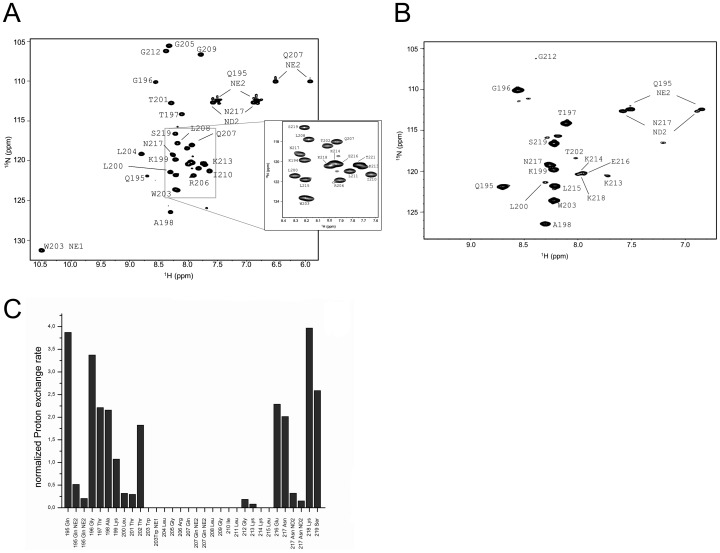
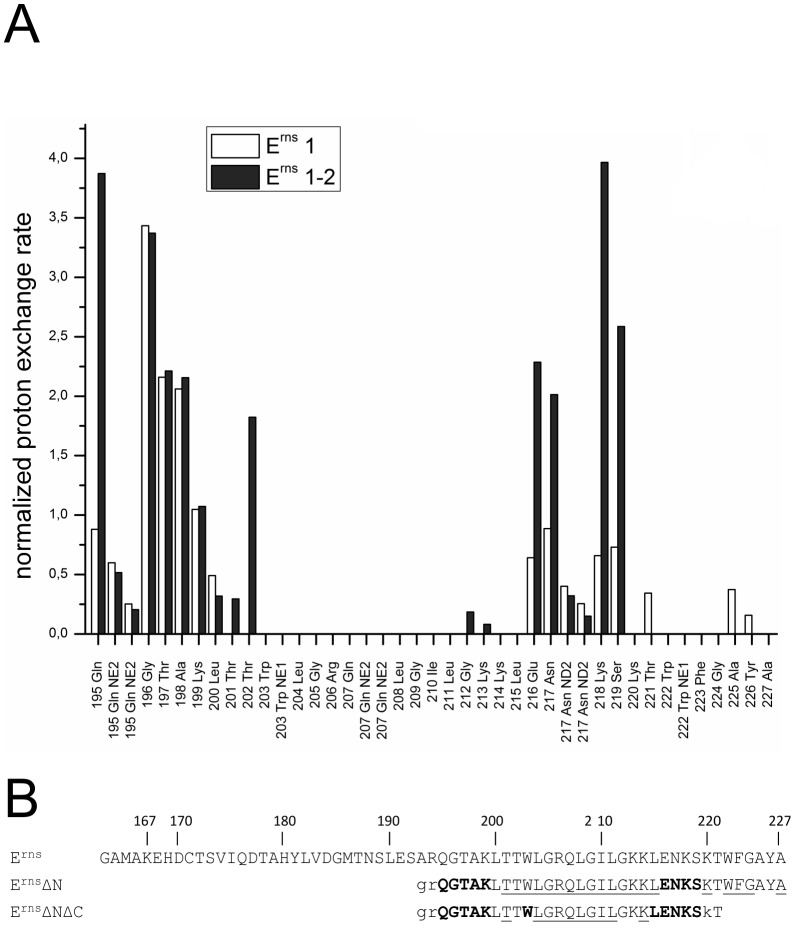
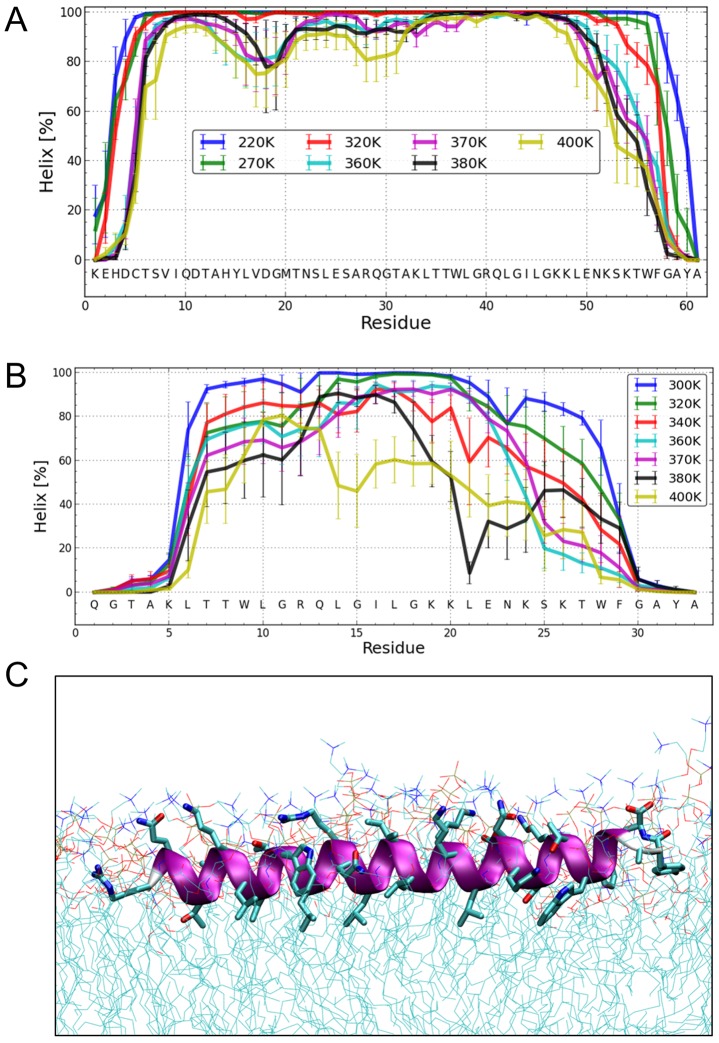
E(rns) is an essential virion glycoprotein with RNase activity that suppresses host cellular innate immune responses upon being partially secreted from the infected cells. Its unusual C-terminus plays multiple roles, as the amphiphilic helix acts as a membrane anchor, as a signal peptidase cleavage site, and as a retention/secretion signal. We analyzed the structure and membrane binding properties of this sequence to gain a better understanding of the underlying mechanisms. CD spectroscopy in different setups, as well as Monte Carlo and molecular dynamics simulations confirmed the helical folding and showed that the helix is accommodated in the amphiphilic region of the lipid bilayer with a slight tilt rather than lying parallel to the surface. This model was confirmed by NMR analyses that also identified a central stretch of 15 residues within the helix that is fully shielded from the aqueous layer, which is C-terminally followed by a putative hairpin structure. These findings explain the strong membrane binding of the protein and provide clues to establishing the E(rns) membrane contact, processing and secretion.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Structures and Functions of Pestivirus Glycoproteins: Not Simply Surface Matters.Viruses. 2015 Jun 29;7(7):3506-29. doi: 10.3390/v7072783. Viruses. 2015. PMID: 26131960 Free PMC article. Review.
-
The Erns Carboxyterminus: Much More Than a Membrane Anchor.Viruses. 2021 Jun 23;13(7):1203. doi: 10.3390/v13071203. Viruses. 2021. PMID: 34201636 Free PMC article.
-
Downstream Sequences Control the Processing of the Pestivirus Erns-E1 Precursor.J Virol. 2020 Dec 9;95(1):e01905-20. doi: 10.1128/JVI.01905-20. Print 2020 Dec 9. J Virol. 2020. PMID: 33028718 Free PMC article.
-
Charged Residues in the Membrane Anchor of the Pestiviral Erns Protein Are Important for Processing and Secretion of Erns and Recovery of Infectious Viruses.Viruses. 2021 Mar 10;13(3):444. doi: 10.3390/v13030444. Viruses. 2021. PMID: 33801849 Free PMC article.
-
A new type of intracellular retention signal identified in a pestivirus structural glycoprotein.FASEB J. 2012 Aug;26(8):3292-305. doi: 10.1096/fj.12-207191. Epub 2012 May 1. FASEB J. 2012. PMID: 22549508
Cited by
-
Host Cell Receptors Implicated in the Cellular Tropism of BVDV.Viruses. 2022 Oct 20;14(10):2302. doi: 10.3390/v14102302. Viruses. 2022. PMID: 36298858 Free PMC article. Review.
-
Host-derived apolipoproteins play comparable roles with viral secretory proteins Erns and NS1 in the infectious particle formation of Flaviviridae.PLoS Pathog. 2017 Jun 23;13(6):e1006475. doi: 10.1371/journal.ppat.1006475. eCollection 2017 Jun. PLoS Pathog. 2017. PMID: 28644867 Free PMC article.
-
Structures and Functions of Pestivirus Glycoproteins: Not Simply Surface Matters.Viruses. 2015 Jun 29;7(7):3506-29. doi: 10.3390/v7072783. Viruses. 2015. PMID: 26131960 Free PMC article. Review.
-
The Molecular Basis for Erns Dimerization in Classical Swine Fever Virus.Viruses. 2021 Nov 2;13(11):2204. doi: 10.3390/v13112204. Viruses. 2021. PMID: 34835010 Free PMC article.
-
The Erns Carboxyterminus: Much More Than a Membrane Anchor.Viruses. 2021 Jun 23;13(7):1203. doi: 10.3390/v13071203. Viruses. 2021. PMID: 34201636 Free PMC article.
References
-
- Lindenbach BD, Thiel HJ, Rice CM (2007) Flaviviridae: The Viruses and Their Replication. In: Knipe DM, Howley PM, editors. Fields Virology. Philadelphia, New York: Lippincott - Raven Publishers. pp. 1101–1152.
-
- Moennig V, Plagemann PG (1992) The pestiviruses. Advances in virus research 41: 53–98. - PubMed
-
- Thiel HJ, Plagemann PGW, Moennig V (1996) Pestiviruses. In: Fields BN, Knipe DM, Howley PM, editors. Fields Virology. Philadelphia, New York: Lippincott - Raven Publishers. pp. 1059–1073.
-
- Seago J, Hilton L, Reid E, Doceul V, Jeyatheesan J, et al. (2007) The Npro product of classical swine fever virus and bovine viral diarrhea virus uses a conserved mechanism to target interferon regulatory factor-3. The Journal of general virology 88: 3002–3006. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
