

Myc and max genome-wide binding sites analysis links the Myc regulatory network with the polycomb and the core pluripotency networks in mouse embryonic stem cells
- PMID: 24586446
- PMCID: PMC3931652
- DOI: 10.1371/journal.pone.0088933
Myc and max genome-wide binding sites analysis links the Myc regulatory network with the polycomb and the core pluripotency networks in mouse embryonic stem cells
Abstract
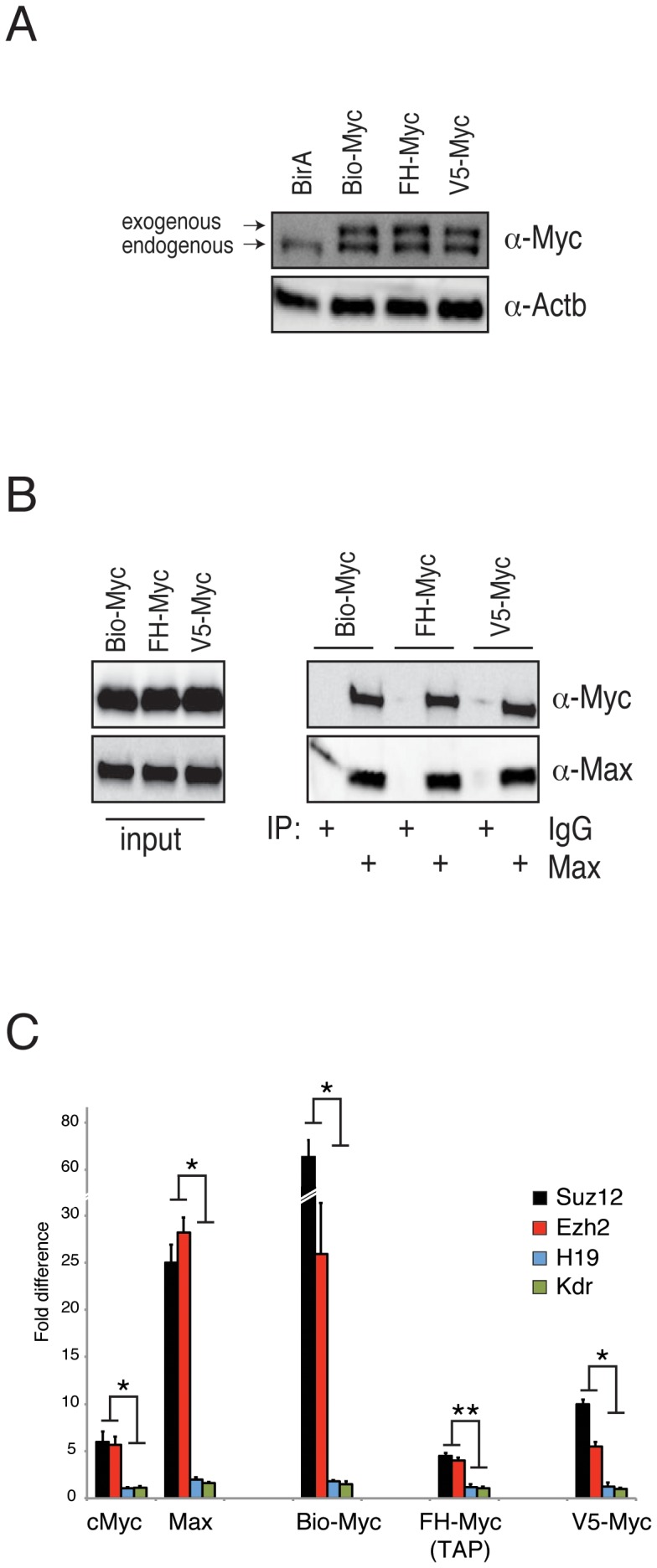
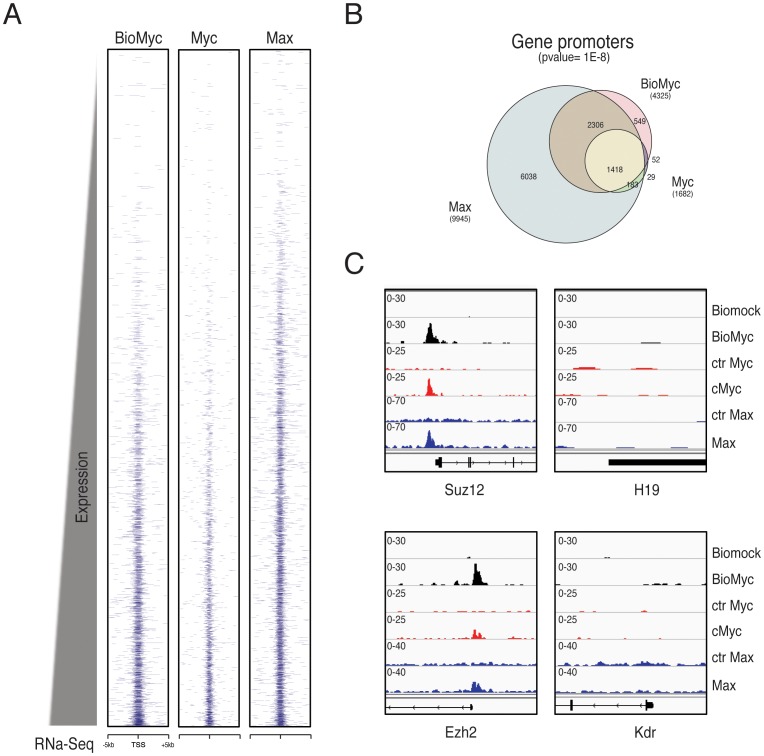
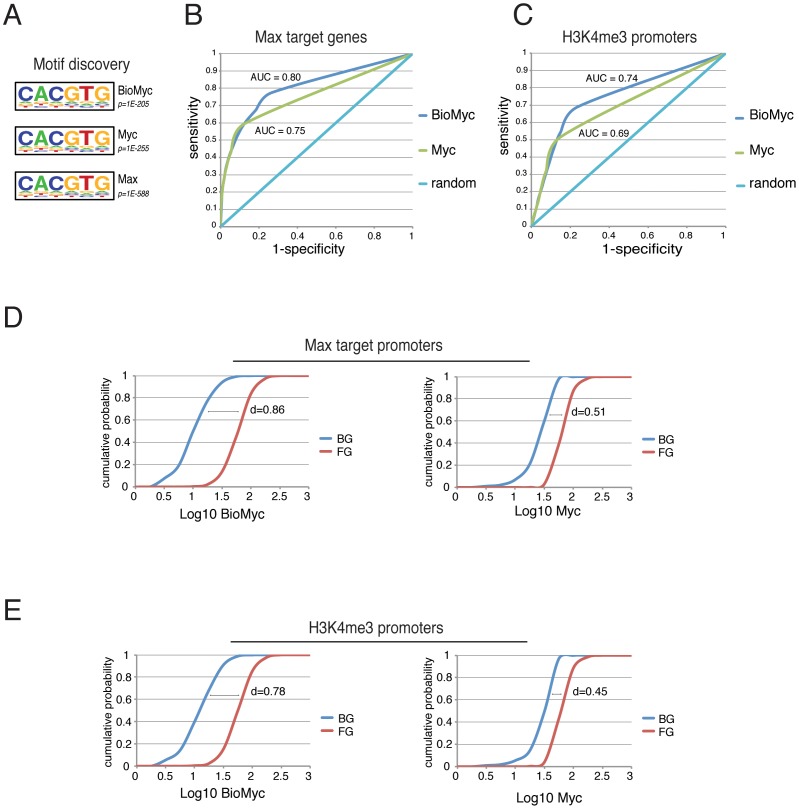
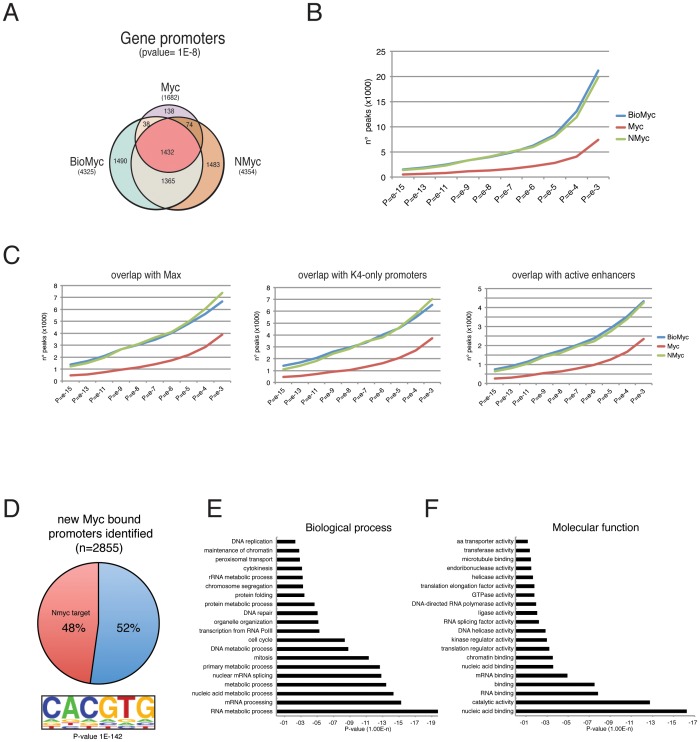
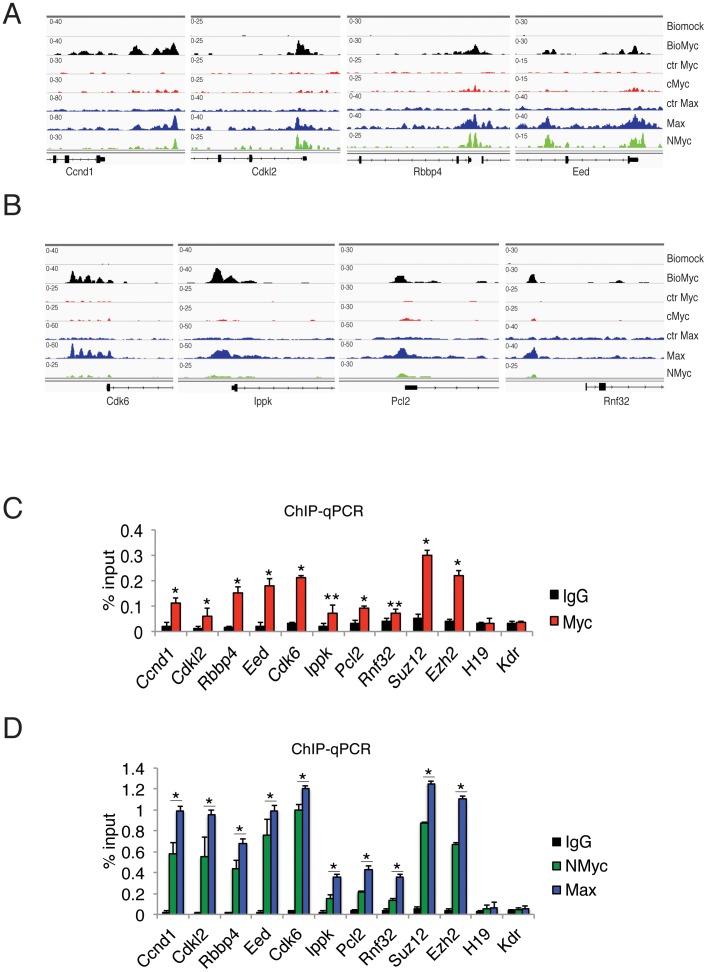
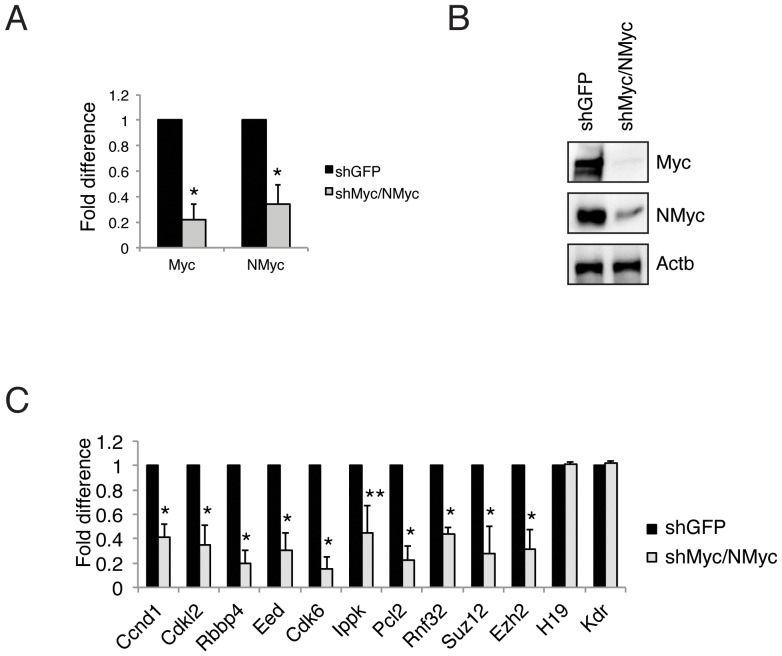
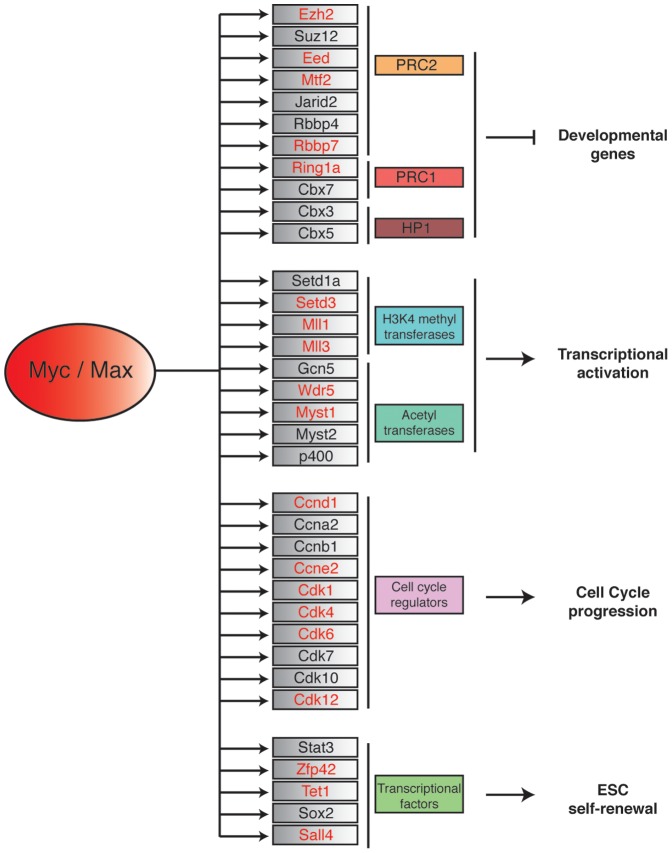
Myc is a master transcription factor that has been demonstrated to be required for embryonic stem cell (ESC) pluripotency, self-renewal, and inhibition of differentiation. Although recent works have identified several Myc-targets in ESCs, the list of Myc binding sites is largely incomplete due to the low sensitivity and specificity of the antibodies available. To systematically identify Myc binding sites in mouse ESCs, we used a stringent streptavidin-based genome-wide chromatin immunoprecipitation (ChIP-Seq) approach with biotin-tagged Myc (Bio-Myc) as well as a ChIP-Seq of the Myc binding partner Max. This analysis identified 4325 Myc binding sites, of which 2885 were newly identified. The identified sites overlap with more than 85% of the Max binding sites and are enriched for H3K4me3-positive promoters and active enhancers. Remarkably, this analysis unveils that Myc/Max regulates chromatin modifiers and transcriptional regulators involved in stem cell self-renewal linking the Myc-centered network with the Polycomb and the Core networks. These results provide insights into the contribution of Myc and Max in maintaining stem cell self-renewal and keeping these cells in an undifferentiated state.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
