

The activated SA and JA signaling pathways have an influence on flg22-triggered oxidative burst and callose deposition
- PMID: 24586453
- PMCID: PMC3934882
- DOI: 10.1371/journal.pone.0088951
The activated SA and JA signaling pathways have an influence on flg22-triggered oxidative burst and callose deposition
Abstract
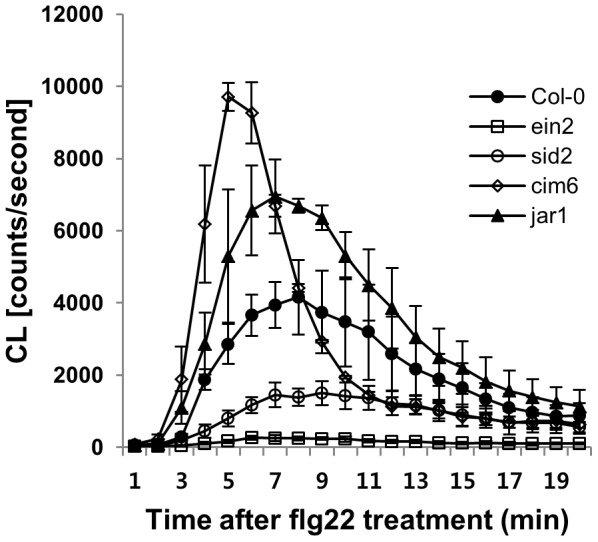
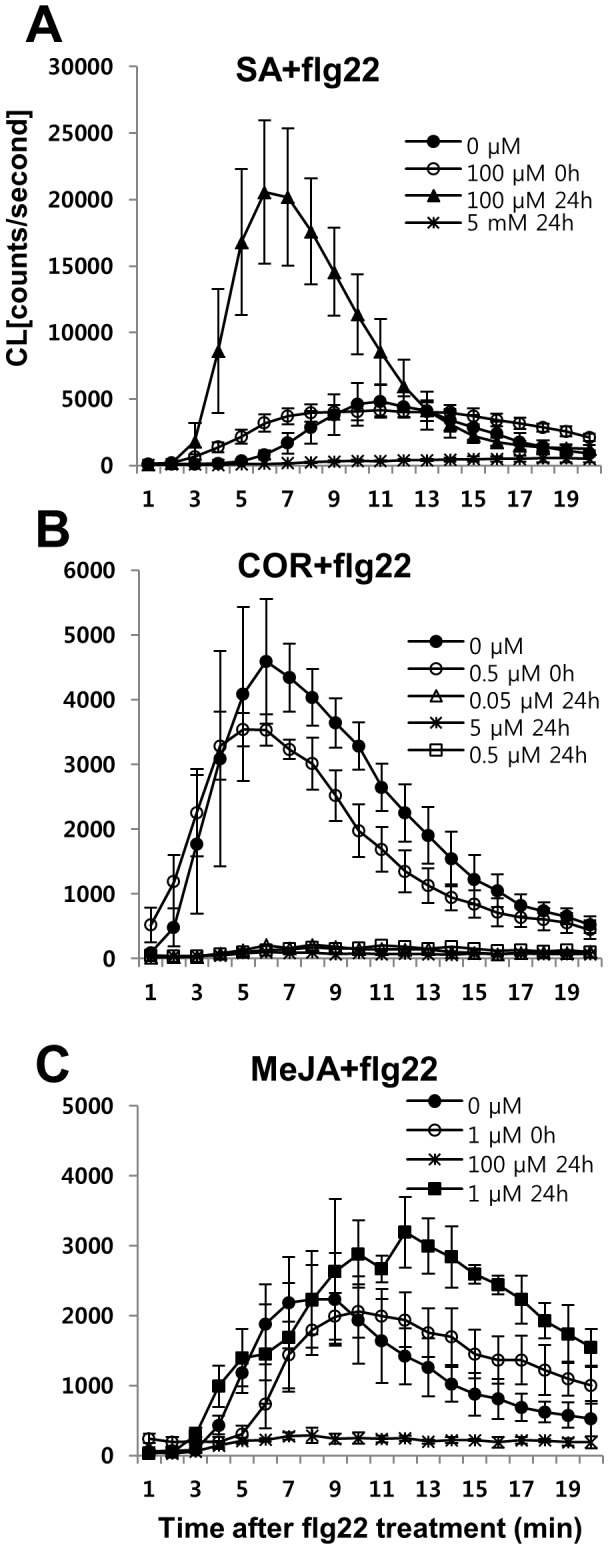
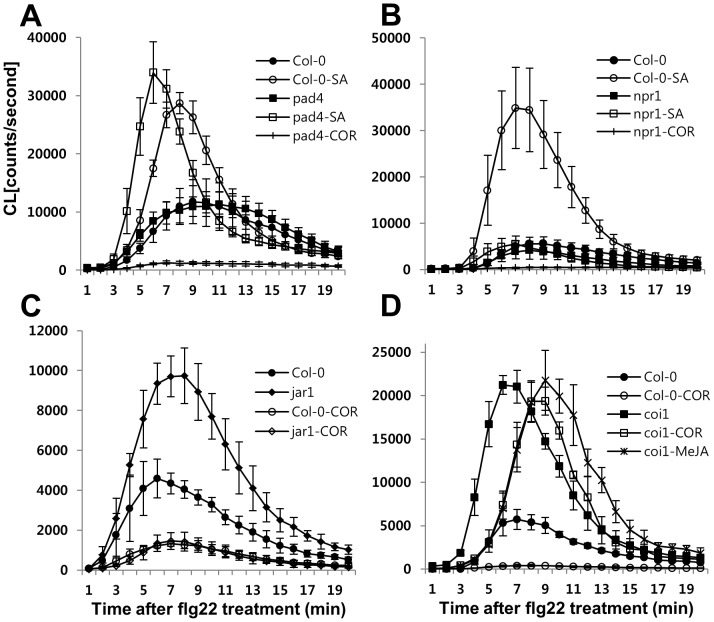
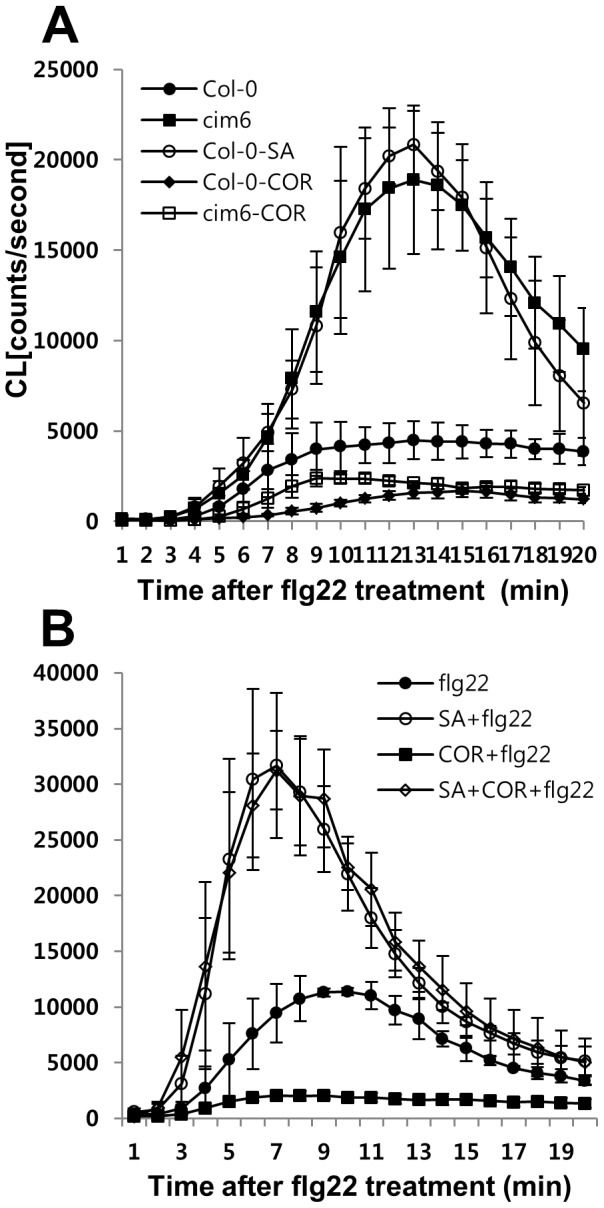
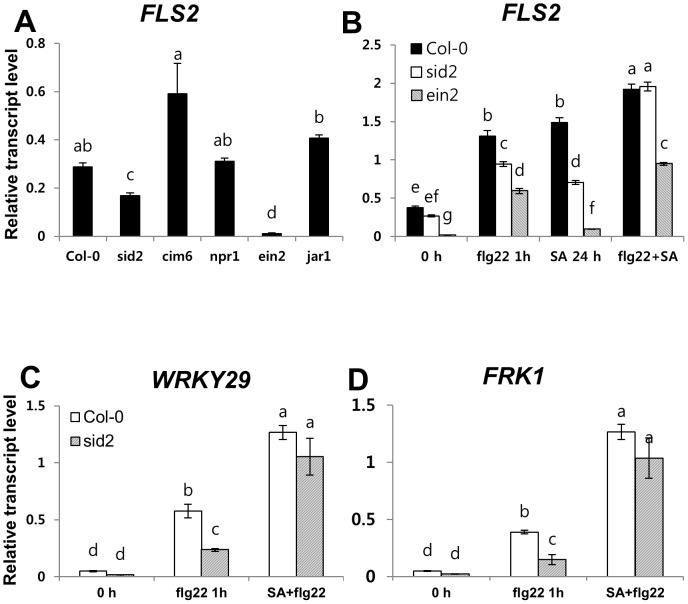
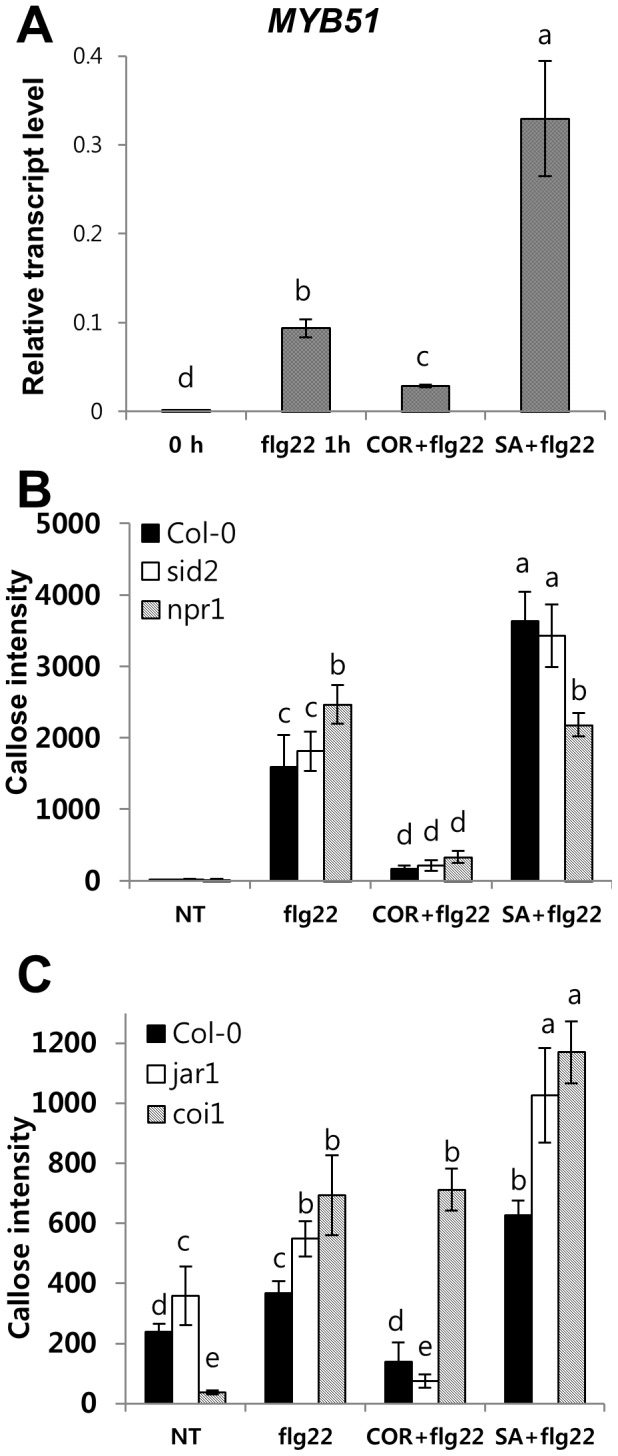
The first line of defense in plants against pathogens is induced by the recognition of microbe-associated molecular patterns (MAMP). Perception of bacterial flagellin (flg22) by the pattern recognition receptor flagellin-sensing 2 (FLS2) is the best characterized MAMP response, although the underlying molecular mechanisms are not fully understood. Here we studied the relationship between salicylic acid (SA) or jasmonic acid (JA) signaling and FLS2-mediated signaling by monitoring flg22-triggered responses in known SA or JA related mutants of Arabidopsis thaliana (L.) Heynh. The sid2 mutant, impaired in SA biosynthesis, had less basal FLS2 mRNA accumulation than the wild type, which correlated with suppression of early flg22 responses such as ROS production and induction of marker genes, WRKY29 and FRK1. The JA-signaling mutants, jar1 and coi1, exhibited an enhanced flg22-triggered oxidative burst and more callose accumulation than the wild type, and pretreatment with SA or coronatine (COR), a structural mimic of JA-isoleucine, altered these flg22-induced responses. Nonexpressor of pathogenesis-related genes 1 (NPR1) acted downstream of SID2 and required SA-dependent priming for the enhanced flg22-triggered oxidative burst and callose deposition. Activation of JA signaling by COR pretreatment suppressed the flg22-triggered oxidative burst and callose accumulation in a coronatine insensitive 1 (COI1) dependent manner. COR had a negative effect on flg22 responses but only the flg22-triggered oxidative burst depended on SA-JA/COR signaling antagonism. Thus the activated SA and JA signaling pathways have an influence on flg22-triggered oxidative burst and callose deposition. These results may explain how SA and JA signaling are cross talked for regulation of flg22-triggered responses.
Conflict of interest statement
Figures
References
-
- Jones JD, Dangl JL (2006) The plant immune system. Nature 444: 323–329. - PubMed
-
- Felix G, Duran JD, Volko S, Boller T (1999) Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J 18: 265–276. - PubMed
-
- Gomez-Gomez L, Boller T (2002) Flagellin perception: a paradigm for innate immunity. Trends Plant Sci 7: 251–256. - PubMed
-
- Gomez-Gomez L, Boller T (2000) FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol Cell 5: 1003–1011. - PubMed
-
- Gomez-Gomez L, Felix G, Boller T (1999) A single locus determines sensitivity to bacterial flagellin in Arabidopsis thaliana. Plant J 18: 277–284. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
