

Postsynaptic odorant concentration dependent inhibition controls temporal properties of spike responses of projection neurons in the moth antennal lobe
- PMID: 24586546
- PMCID: PMC3929629
- DOI: 10.1371/journal.pone.0089132
Postsynaptic odorant concentration dependent inhibition controls temporal properties of spike responses of projection neurons in the moth antennal lobe
Abstract
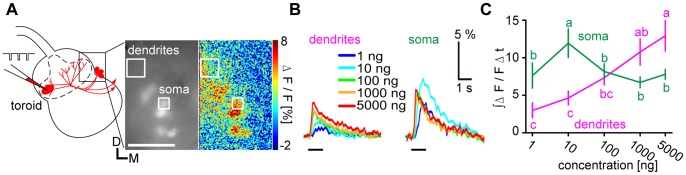
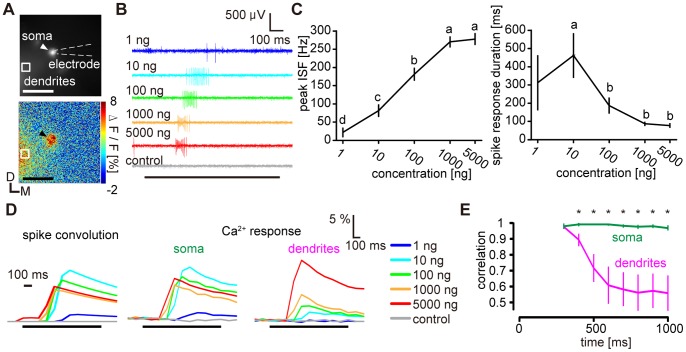
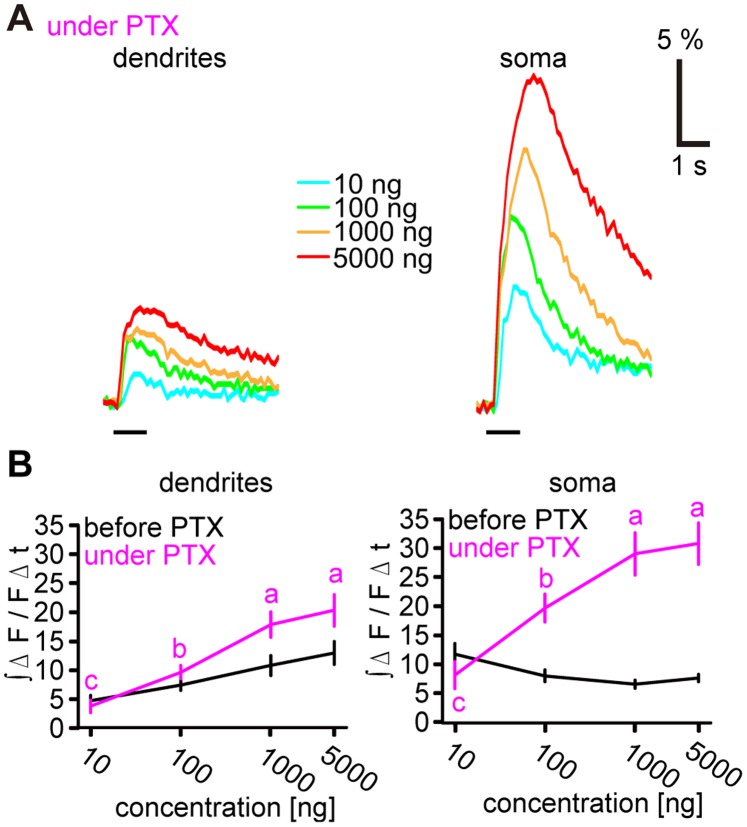
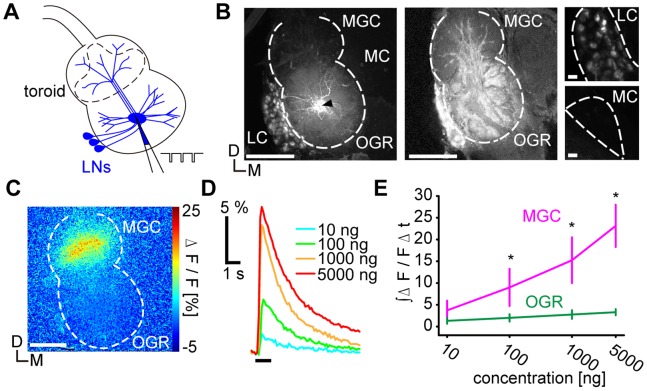
Although odorant concentration-response characteristics of olfactory neurons have been widely investigated in a variety of animal species, the effect of odorant concentration on neural processing at circuit level is still poorly understood. Using calcium imaging in the silkmoth (Bombyx mori) pheromone processing circuit of the antennal lobe (AL), we studied the effect of odorant concentration on second-order projection neuron (PN) responses. While PN calcium responses of dendrites showed monotonic increases with odorant concentration, calcium responses of somata showed decreased responses at higher odorant concentrations due to postsynaptic inhibition. Simultaneous calcium imaging and electrophysiology revealed that calcium responses of PN somata but not dendrites reflect spiking activity. Inhibition shortened spike response duration rather than decreasing peak instantaneous spike frequency (ISF). Local interneurons (LNs) that were specifically activated at high odorant concentrations at which PN responses were suppressed are the putative source of inhibition. Our results imply the existence of an intraglomerular mechanism that preserves time resolution in olfactory processing over a wide odorant concentration range.
Conflict of interest statement
Figures
References
-
- Hallem EA, Dahanukar A, Carlson JR (2006) Insect odor and taste receptors. Annu Rev Entomol 51: 113–135. - PubMed
-
- Wang JW, Wong AM, Flores J, Vosshall LB, Axel R (2003) Two photon calcium imaging reveals an odor evoked map of activity in the fly brain. Cell 112: 271–282. - PubMed
-
- Friedrich RW, Korsching SI (1997) Combinatorial and chemotopic odorant coding in the zebrafish olfactory bulb visualized by optical imaging. Neuron 18: 737–752. - PubMed
-
- Imamura K, Mataga N, Mori K (1992) Coding of odor molecules by mitral tufted cells in rabbit olfactory bulb. I. Aliphatic compounds. J Neurophysiol 68: 1986–2002. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
