

Inhibition of post-transcriptional RNA processing by CDK inhibitors and its implication in anti-viral therapy
- PMID: 24586613
- PMCID: PMC3931720
- DOI: 10.1371/journal.pone.0089228
Inhibition of post-transcriptional RNA processing by CDK inhibitors and its implication in anti-viral therapy
Abstract
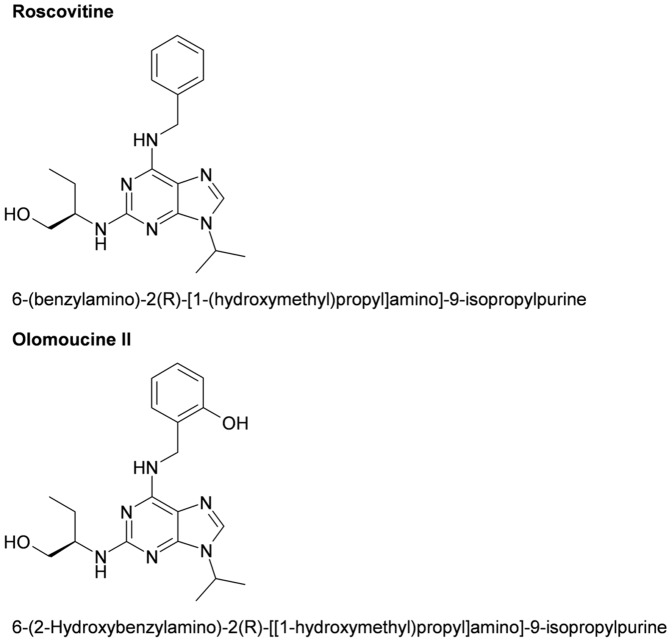
Cyclin-dependent kinases (CDKs) are key regulators of the cell cycle and RNA polymerase II mediated transcription. Several pharmacological CDK inhibitors are currently in clinical trials as potential cancer therapeutics and some of them also exhibit antiviral effects. Olomoucine II and roscovitine, purine-based inhibitors of CDKs, were described as effective antiviral agents that inhibit replication of a broad range of wild type human viruses. Olomoucine II and roscovitine show high selectivity for CDK7 and CDK9, with important functions in the regulation of RNA polymerase II transcription. RNA polymerase II is necessary for viral transcription and following replication in cells. We analyzed the effect of inhibition of CDKs by olomoucine II on gene expression from viral promoters and compared its effect to widely-used roscovitine. We found that both roscovitine and olomoucine II blocked the phosphorylation of RNA polymerase II C-terminal domain. However the repression of genes regulated by viral promoters was strongly dependent on gene localization. Both roscovitine and olomoucine II inhibited expression only when the viral promoter was not integrated into chromosomal DNA. In contrast, treatment of cells with genome-integrated viral promoters increased their expression even though there was decreased phosphorylation of the C-terminal domain of RNA polymerase II. To define the mechanism responsible for decreased gene expression after pharmacological CDK inhibitor treatment, the level of mRNA transcription from extrachromosomal DNA was determined. Interestingly, our results showed that inhibition of RNA polymerase II C-terminal domain phosphorylation increased the number of transcribed mRNAs. However, some of these mRNAs were truncated and lacked polyadenylation, which resulted in decreased translation. These results suggest that phosphorylation of RNA polymerase II C-terminal domain is critical for linking transcription and posttrancriptional processing of mRNA expressed from extrachromosomal DNA.
Conflict of interest statement
Figures






Similar articles
-
Inhibition of the cyclin-dependent kinases at the beginning of human cytomegalovirus infection specifically alters the levels and localization of the RNA polymerase II carboxyl-terminal domain kinases cdk9 and cdk7 at the viral transcriptosome.J Virol. 2008 Jan;82(1):394-407. doi: 10.1128/JVI.01681-07. Epub 2007 Oct 17. J Virol. 2008. PMID: 17942543 Free PMC article.
-
Functional p53 in cells contributes to the anticancer effect of the cyclin-dependent kinase inhibitor roscovitine.J Cell Biochem. 2009 Jun 1;107(3):428-37. doi: 10.1002/jcb.22139. J Cell Biochem. 2009. PMID: 19308936
-
Requirement for cellular cyclin-dependent kinases in herpes simplex virus replication and transcription.J Virol. 1998 Jul;72(7):5626-37. doi: 10.1128/JVI.72.7.5626-5637.1998. J Virol. 1998. PMID: 9621021 Free PMC article.
-
Potential use of pharmacological cyclin-dependent kinase inhibitors as anti-HIV therapeutics.Curr Pharm Des. 2006;12(16):1949-61. doi: 10.2174/138161206777442083. Curr Pharm Des. 2006. PMID: 16787240 Review.
-
Dual action of the inhibitors of cyclin-dependent kinases: targeting of the cell-cycle progression and activation of wild-type p53 protein.Expert Opin Investig Drugs. 2006 Jan;15(1):23-38. doi: 10.1517/13543784.15.1.23. Expert Opin Investig Drugs. 2006. PMID: 16370931 Review.
Cited by
-
Cyclin-dependent kinase activity is required for type I interferon production.Proc Natl Acad Sci U S A. 2018 Mar 27;115(13):E2950-E2959. doi: 10.1073/pnas.1720431115. Epub 2018 Mar 5. Proc Natl Acad Sci U S A. 2018. PMID: 29507205 Free PMC article.
-
Improving the Understanding of Pathogenesis of Human Papillomavirus 16 via Mapping Protein-Protein Interaction Network.Biomed Res Int. 2015;2015:890381. doi: 10.1155/2015/890381. Epub 2015 Apr 15. Biomed Res Int. 2015. PMID: 25961044 Free PMC article.
-
Temporal Proteomic Analysis of BK Polyomavirus Infection Reveals Virus-Induced G2 Arrest and Highly Effective Evasion of Innate Immune Sensing.J Virol. 2019 Jul 30;93(16):e00595-19. doi: 10.1128/JVI.00595-19. Print 2019 Aug 15. J Virol. 2019. PMID: 31142673 Free PMC article.
-
Repurposing of CDK Inhibitors as Host Targeting Antivirals: A Mini- Review.Mini Rev Med Chem. 2025;25(3):178-189. doi: 10.2174/0113895575311618240820103549. Mini Rev Med Chem. 2025. PMID: 39185650 Review.
-
Studies on quantitative phosphopeptide analysis by matrix-assisted laser desorption/ionization mass spectrometry without label, chromatography or calibration curves.Rapid Commun Mass Spectrom. 2014 Dec 30;28(24):2681-9. doi: 10.1002/rcm.7063. Rapid Commun Mass Spectrom. 2014. PMID: 25380489 Free PMC article.
References
-
- Knockaert M, Greengard P, Meijer L (2002) Pharmacological inhibitors of cyclin-dependent kinases. Trends Pharmacol Sci 23: 417–425. - PubMed
-
- Arris CE, Boyle FT, Calvert AH, Curtin NJ, Endicott JA, et al. (2000) Identification of novel purine and pyrimidine cyclin-dependent kinase inhibitors with distinct molecular interactions and tumor cell growth inhibition profiles. J Med Chem 43: 2797–2804. - PubMed
-
- Havlicek L, Hanus J, Vesely J, Leclerc S, Meijer L, et al. (1997) Cytokinin-derived cyclin-dependent kinase inhibitors: synthesis and cdc2 inhibitory activity of olomoucine and related compounds. J Med Chem 40: 408–412. - PubMed
-
- Krystof V, Lenobel R, Havlicek L, Kuzma M, Strnad M (2002) Synthesis and biological activity of olomoucine II. Bioorg Med Chem Lett 12: 3283–3286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
