

The ubiquitin ligase ASB4 promotes trophoblast differentiation through the degradation of ID2
- PMID: 24586788
- PMCID: PMC3931756
- DOI: 10.1371/journal.pone.0089451
The ubiquitin ligase ASB4 promotes trophoblast differentiation through the degradation of ID2
Abstract
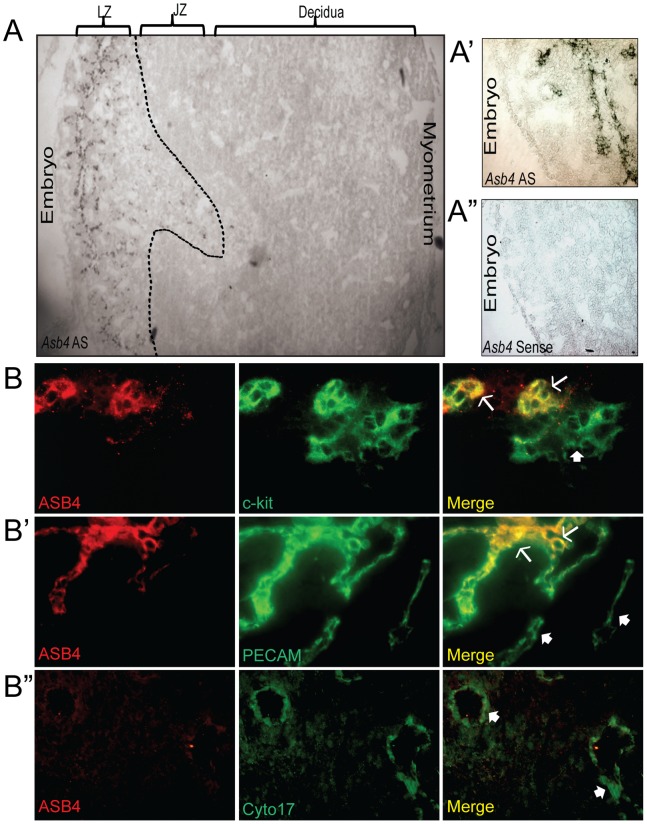
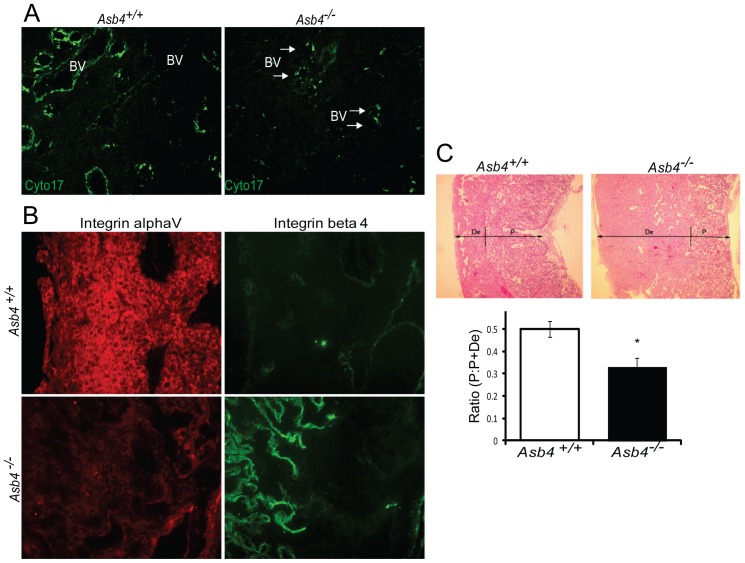
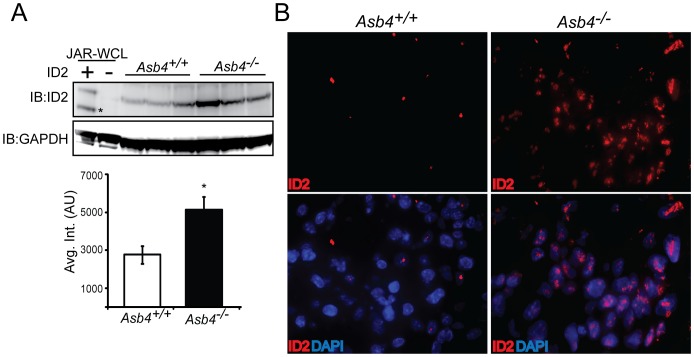
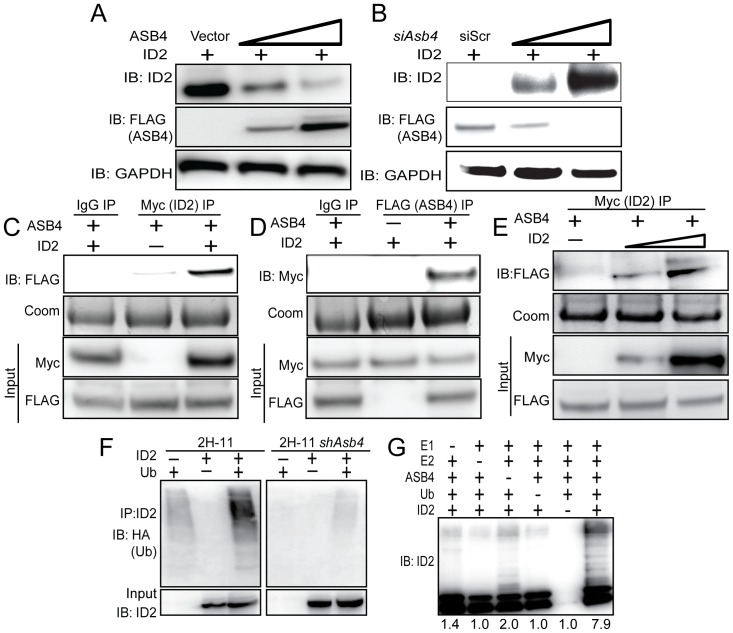
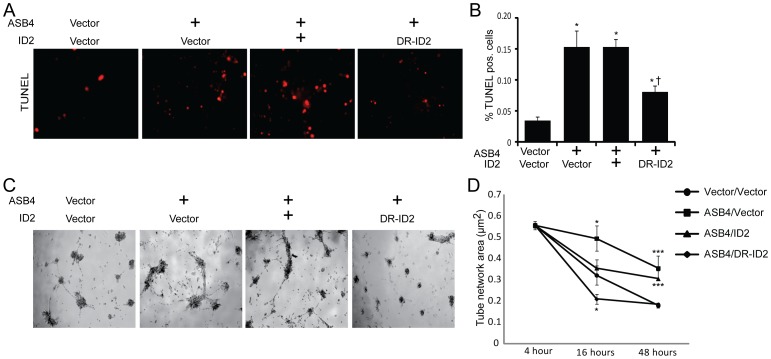
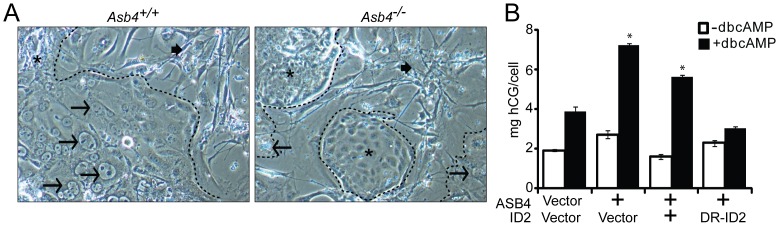
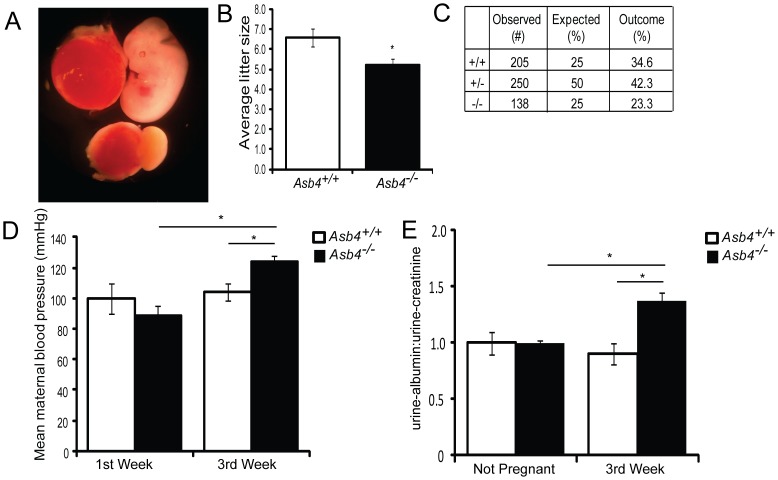
Vascularization of the placenta is a critical developmental process that ensures fetal viability. Although the vascular health of the placenta affects both maternal and fetal well being, relatively little is known about the early stages of placental vascular development. The ubiquitin ligase Ankyrin repeat, SOCS box-containing 4 (ASB4) promotes embryonic stem cell differentiation to vascular lineages and is highly expressed early in placental development. The transcriptional regulator Inhibitor of DNA binding 2 (ID2) negatively regulates vascular differentiation during development and is a target of many ubiquitin ligases. Due to their overlapping spatiotemporal expression pattern in the placenta and contrasting effects on vascular differentiation, we investigated whether ASB4 regulates ID2 through its ligase activity in the placenta and whether this activity mediates vascular differentiation. In mouse placentas, ASB4 expression is restricted to a subset of cells that express both stem cell and endothelial markers. Placentas that lack Asb4 display immature vascular patterning and retain expression of placental progenitor markers, including ID2 expression. Using JAR placental cells, we determined that ASB4 ubiquitinates and represses ID2 expression in a proteasome-dependent fashion. Expression of ASB4 in JAR cells and primary isolated trophoblast stem cells promotes the expression of differentiation markers. In functional endothelial co-culture assays, JAR cells ectopically expressing ASB4 increased endothelial cell turnover and stabilized endothelial tube formation, both of which are hallmarks of vascular differentiation within the placenta. Co-transfection of a degradation-resistant Id2 mutant with Asb4 inhibits both differentiation and functional responses. Lastly, deletion of Asb4 in mice induces a pathology that phenocopies human pre-eclampsia, including hypertension and proteinuria in late-stage pregnant females. These results indicate that ASB4 mediates vascular differentiation in the placenta via its degradation of ID2.
Conflict of interest statement
Figures
Similar articles
-
Insulin Elevates ID2 Expression in Trophoblasts and Aggravates Preeclampsia in Obese ASB4-Null Mice.Int J Mol Sci. 2023 Jan 21;24(3):2149. doi: 10.3390/ijms24032149. Int J Mol Sci. 2023. PMID: 36768469 Free PMC article.
-
ASB4 is a hydroxylation substrate of FIH and promotes vascular differentiation via an oxygen-dependent mechanism.Mol Cell Biol. 2007 Sep;27(18):6407-19. doi: 10.1128/MCB.00511-07. Epub 2007 Jul 16. Mol Cell Biol. 2007. PMID: 17636018 Free PMC article.
-
The Transcription Factor OVOL2 Represses ID2 and Drives Differentiation of Trophoblast Stem Cells and Placental Development in Mice.Cells. 2020 Mar 31;9(4):840. doi: 10.3390/cells9040840. Cells. 2020. PMID: 32244352 Free PMC article.
-
The Roles of Obesity and ASB4 in Preeclampsia Pathogenesis.Int J Mol Sci. 2024 Aug 20;25(16):9017. doi: 10.3390/ijms25169017. Int J Mol Sci. 2024. PMID: 39201703 Free PMC article. Review.
-
Placental formation in early pregnancy: how is the centre of the placenta made?Hum Reprod Update. 2018 Nov 1;24(6):750-760. doi: 10.1093/humupd/dmy030. Hum Reprod Update. 2018. PMID: 30257012 Review.
Cited by
-
Pilot Study on the Profiling and Functional Analysis of mRNA, miRNA, and lncRNA in the Skeletal Muscle of Mongolian Horses, Xilingol Horses, and Grassland-Thoroughbreds.Animals (Basel). 2025 Apr 13;15(8):1123. doi: 10.3390/ani15081123. Animals (Basel). 2025. PMID: 40281957 Free PMC article.
-
Vitamin B3 Nicotinamide: A Promising Candidate for Treating Preeclampsia and Improving Fetal Growth.Tohoku J Exp Med. 2018 Mar;244(3):243-248. doi: 10.1620/tjem.244.243. Tohoku J Exp Med. 2018. PMID: 29563389 Free PMC article. Review.
-
The Role of Oxygen Sensors, Hydroxylases, and HIF in Cardiac Function and Disease.Oxid Med Cell Longev. 2015;2015:676893. doi: 10.1155/2015/676893. Epub 2015 Sep 28. Oxid Med Cell Longev. 2015. PMID: 26491535 Free PMC article. Review.
-
E3 Ubiquitin Ligases Neurobiological Mechanisms: Development to Degeneration.Front Mol Neurosci. 2017 May 19;10:151. doi: 10.3389/fnmol.2017.00151. eCollection 2017. Front Mol Neurosci. 2017. PMID: 28579943 Free PMC article. Review.
-
Epigenetics of Skeletal Muscle-Associated Genes in the ASB, LRRC, TMEM, and OSBPL Gene Families.Epigenomes. 2020 Jan 30;4(1):1. doi: 10.3390/epigenomes4010001. Epigenomes. 2020. PMID: 34968235 Free PMC article.
References
-
- Ferguson JE 3rd, Kelley RW, Patterson C (2005) Mechanisms of endothelial differentiation in embryonic vasculogenesis. Arterioscler Thromb Vasc Biol 25: 2246–2254. - PubMed
-
- Drake CJ, Fleming PA (2000) Vasculogenesis in the day 6.5 to 9.5 mouse embryo. Blood 95: 1671–1679. - PubMed
-
- Downs KM (1998) The murine allantois. Curr Top Dev Biol 39: 1–33. - PubMed
-
- Conway SJ, Kruzynska-Frejtag A, Kneer PL, Machnicki M, Koushik SV (2003) What cardiovascular defect does my prenatal mouse mutant have, and why? Genesis 35: 1–21. - PubMed
-
- Kam EP, Gardner L, Loke YW, King A (1999) The role of trophoblast in the physiological change in decidual spiral arteries. Hum Reprod 14: 2131–2138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
