

Novel inhibitors of mitochondrial sn-glycerol 3-phosphate dehydrogenase
- PMID: 24587137
- PMCID: PMC3933693
- DOI: 10.1371/journal.pone.0089938
Novel inhibitors of mitochondrial sn-glycerol 3-phosphate dehydrogenase
Abstract
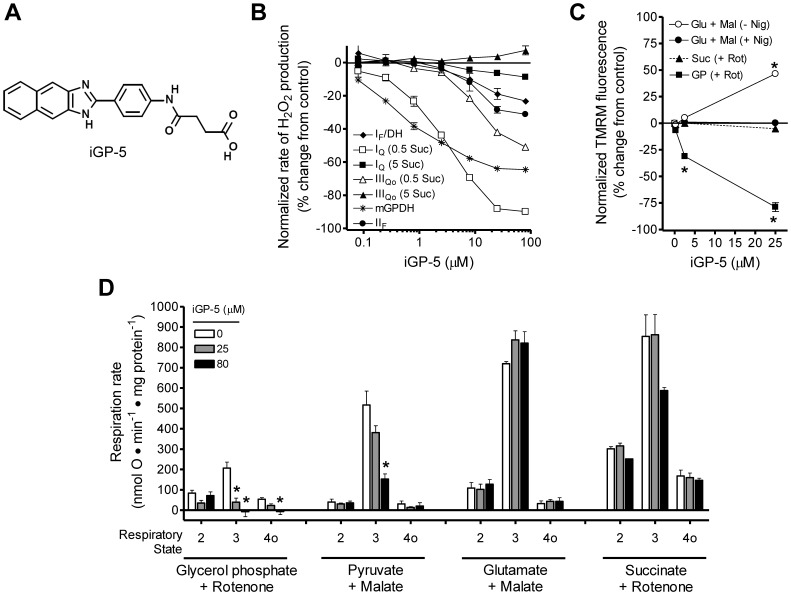
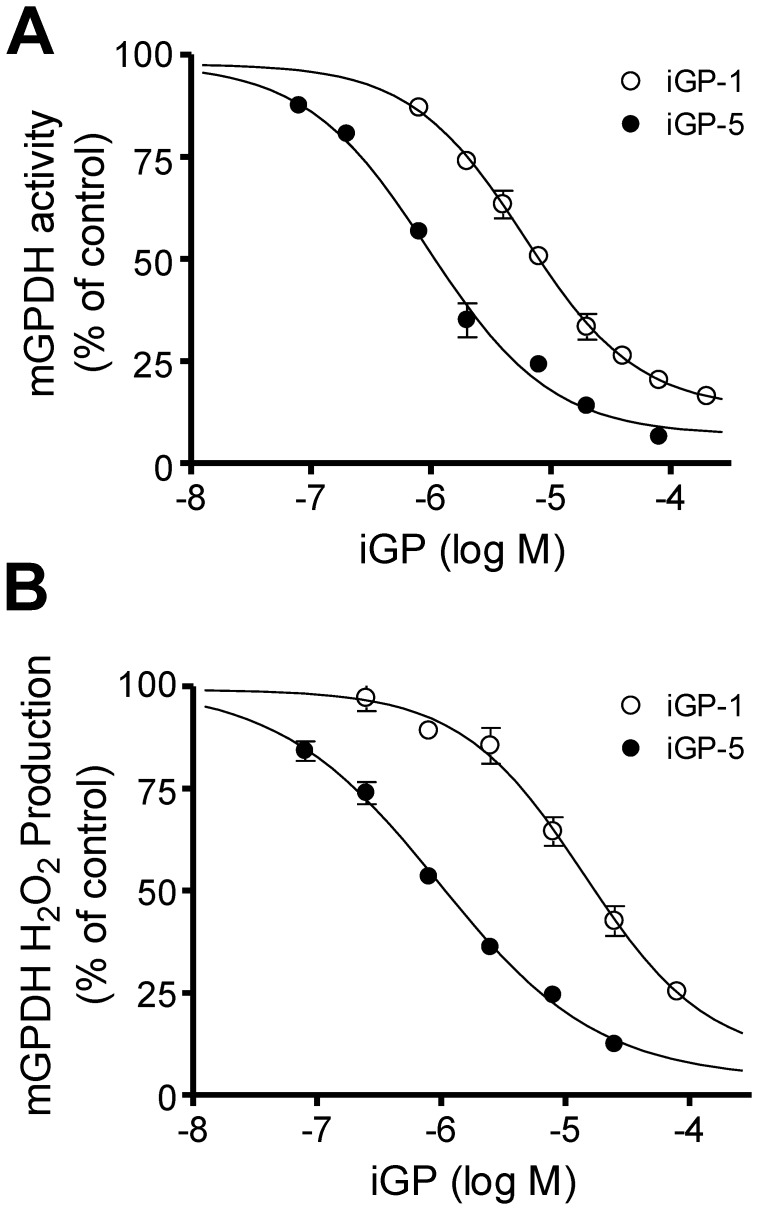
Mitochondrial sn-glycerol 3-phosphate dehydrogenase (mGPDH) is a ubiquinone-linked enzyme in the mitochondrial inner membrane best characterized as part of the glycerol phosphate shuttle that transfers reducing equivalents from cytosolic NADH into the mitochondrial electron transport chain. Despite the widespread expression of mGPDH and the availability of mGPDH-null mice, the physiological role of this enzyme remains poorly defined in many tissues, likely because of compensatory pathways for cytosolic regeneration of NAD⁺ and mechanisms for glycerol phosphate metabolism. Here we describe a novel class of cell-permeant small-molecule inhibitors of mGPDH (iGP) discovered through small-molecule screening. Structure-activity analysis identified a core benzimidazole-phenyl-succinamide structure as being essential to inhibition of mGPDH while modifications to the benzimidazole ring system modulated both potency and off-target effects. Live-cell imaging provided evidence that iGPs penetrate cellular membranes. Two compounds (iGP-1 and iGP-5) were characterized further to determine potency and selectivity and found to be mixed inhibitors with IC₅₀ and K(i) values between ∼1-15 µM. These novel mGPDH inhibitors are unique tools to investigate the role of glycerol 3-phosphate metabolism in both isolated and intact systems.
Conflict of interest statement
Figures








References
-
- Klingenberg M (1970) Localization of the glycerol-phosphate dehydrogenase in the outer phase of the mitochondrial inner membrane. Eur J Biochem 13: 247–252. - PubMed
-
- McKenna MC, Waagepetersen HS, Schousboe A, Sonnewald U (2006) Neuronal and astrocytic shuttle mechanisms for cytosolic-mitochondrial transfer of reducing equivalents: Current evidence and pharmacological tools. Biochem Pharmacol 71: 399–407. - PubMed
-
- Wernette ME, Ochs RS, Lardy HA (1981) Ca2+ stimulation of rat liver mitochondrial glycerophosphate dehydrogenase. J Biol Chem 256: 12767–12771. - PubMed
-
- Brown LJ, MacDonald MJ, Lehn DA, Moran SM (1994) Sequence of rat mitochondrial glycerol-3-phosphate dehydrogenase cDNA. Evidence for EF-hand calcium-binding domains. J Biol Chem 269: 14363–14366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
