

Complexity of central processing in simple and choice multilimb reaction-time tasks
- PMID: 24587371
- PMCID: PMC3938735
- DOI: 10.1371/journal.pone.0090457
Complexity of central processing in simple and choice multilimb reaction-time tasks
Abstract
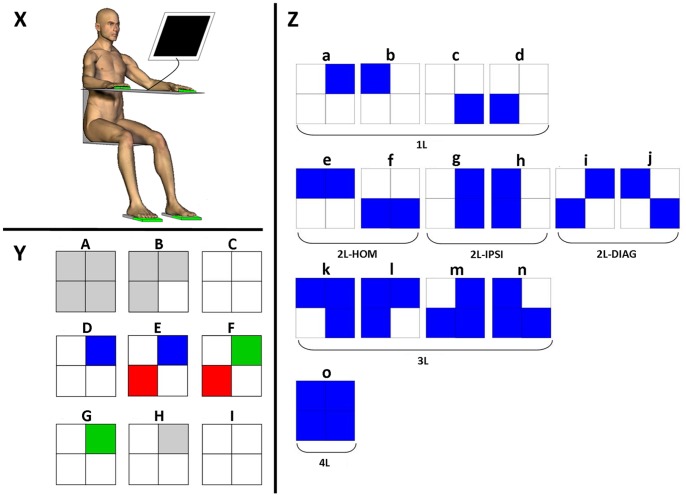
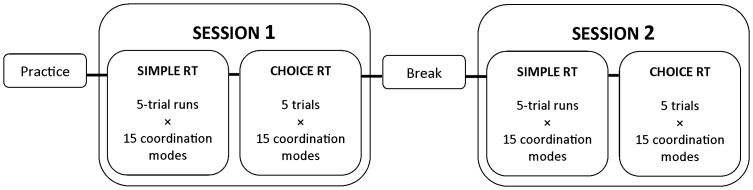
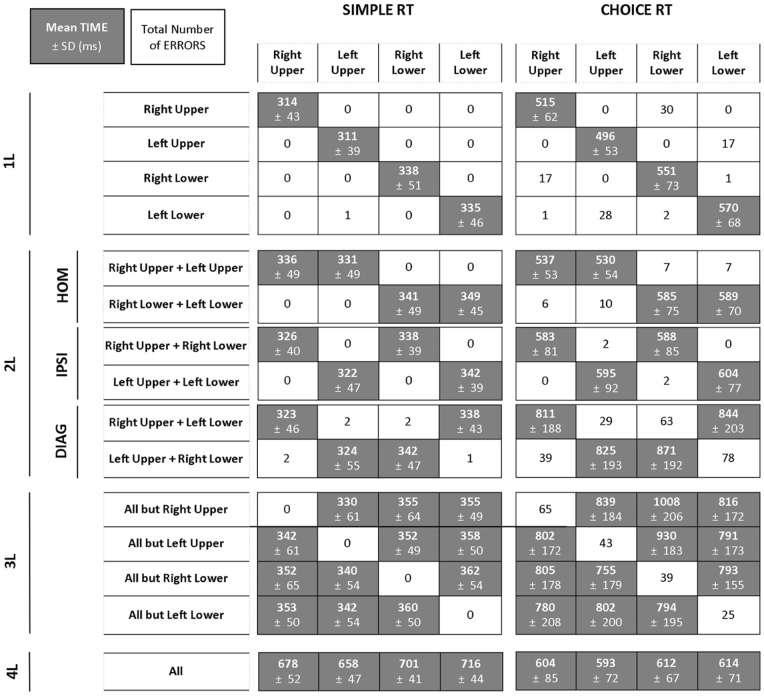
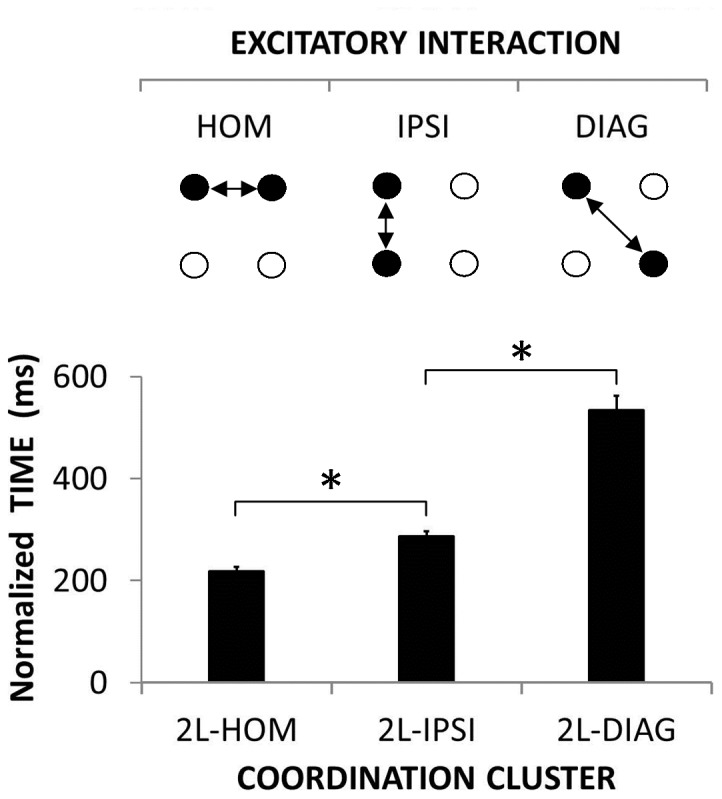
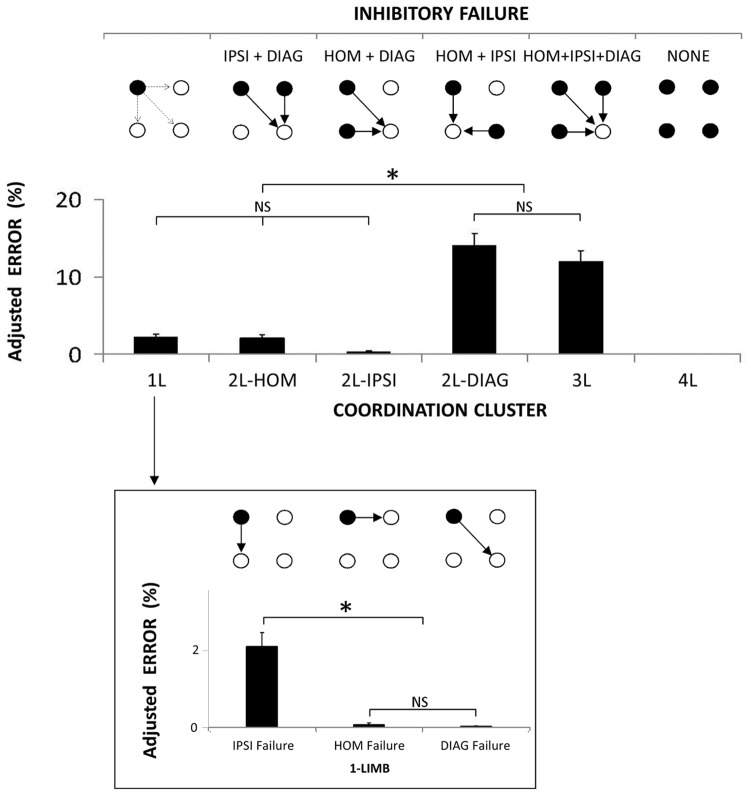
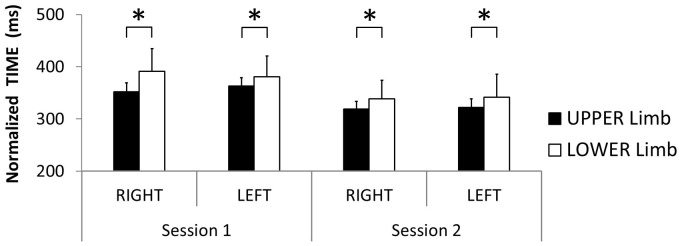
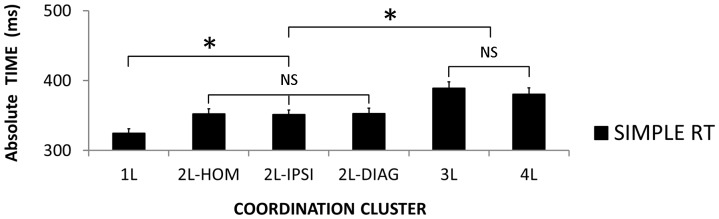
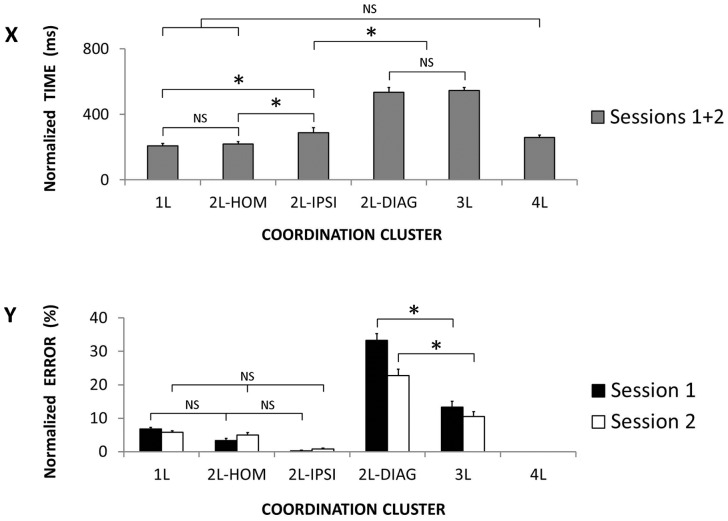
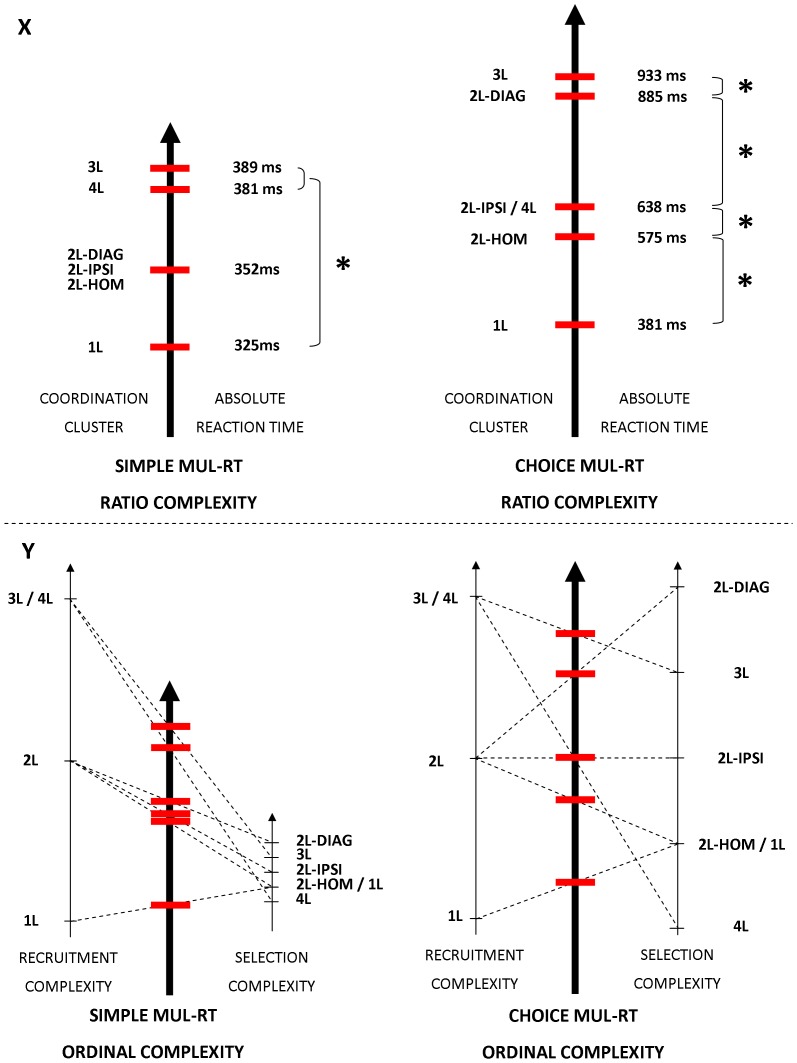
The default mode of the motor system is a coupling between limbs. However, in some movements, a decoupling is required and thus calls for selection and facilitation/inhibition processes. Here, we investigate the relative contribution of recruitment versus selection processes to the overall processing complexity. To this aim we proposed a new multilimb reaction-time task (MUL-RT). Simple, choice and normalized (choice minus simple) RT were analysed together with error rates in thirty-six young adults for 15 coordination modes including all possible configuration of limb recruitment. Simple and normalized RTs were respectively assumed to be indicative of the recruitment and selection processes. Results supported a model of coupling/decoupling interactions respectively reporting weak, intermediate and strong interaction for selecting diagonal, ipsilateral and homologous limbs. Movement laterality (left vs. right) had no effect on selection complexity, whereas selecting upper limbs was less challenging than selecting lower limbs. Results in the different coordination modes suggested that recruitment complexity decreased as follows: 3 limbs = 4 limbs>2 limbs (homologous, ipsilateral and diagonal)>1 limb, and selection complexity as follows: 2 diagonal limbs>3 limbs>2 ipsilateral limbs>1 limb = 2 homologous limbs>4 limbs. Based on these ordinal scales of recruitment and selection complexity, we extrapolated the overall processing complexity of the simple and choice MUL-RT. This method was efficient in reproducing the absolute results we obtained on a ratio scale (ms) and demonstrated that processing complexity in simple RT was mainly governed by the 'recruitment principle' (the more limbs recruited the lower the performance), whereas contributions of recruitment and 'selection principle' (nature of the coordination determines performance) to overall processing complexity were similar in choice RT.
Conflict of interest statement
Figures

References
-
- Jensen AR (2006) Clocking the mind: mental chronometry and individual differences. Oxford: Elsevier. 287p.
-
- Schmidt RA, Lee TD (2005) Motor control and learning: a behavioral emphasis. Champaign: Human Kinetics. 537 p.
-
- Jensen AR (1993) Why is reaction time correlated with psychometric g? Curr Dir Psychol Sci 2: 53–56.
-
- Donders FC (1969) On the speed of mental processes. Acta Psychol (Amst) 30: 412–431. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
