

Parasite-derived microRNAs in host serum as novel biomarkers of helminth infection
- PMID: 24587461
- PMCID: PMC3930507
- DOI: 10.1371/journal.pntd.0002701
Parasite-derived microRNAs in host serum as novel biomarkers of helminth infection
Abstract
Background: MicroRNAs (miRNAs) are a class of short non-coding RNA that play important roles in disease processes in animals and are present in a highly stable cell-free form in body fluids. Here, we examine the capacity of host and parasite miRNAs to serve as tissue or serum biomarkers of Schistosoma mansoni infection.
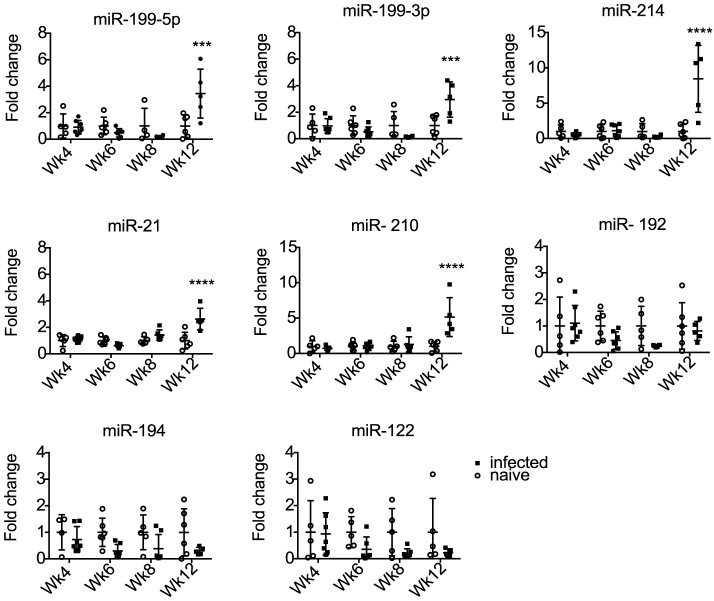
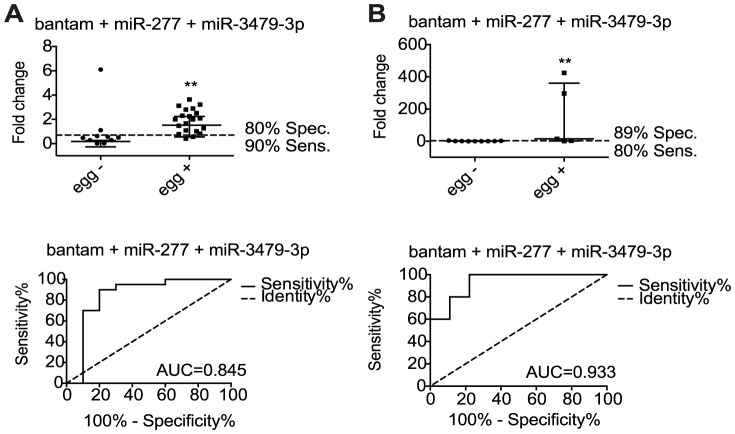
Methods/principal findings: We used Exiqon miRNA microarrays to profile miRNA expression in the livers of mice infected with S. mansoni at 7 weeks post-infection. Thirty-three mouse miRNAs were differentially expressed in infected compared to naïve mice (>2 fold change, p<0.05) including miR-199a-3p, miR-199a-5p, miR-214 and miR-21, which have previously been associated with liver fibrosis in other settings. Five of the mouse miRNAs were also significantly elevated in serum by twelve weeks post-infection. Sequencing of small RNAs from serum confirmed the presence of these miRNAs and further revealed eleven parasite-derived miRNAs that were detectable by eight weeks post infection. Analysis of host and parasite miRNA abundance by qRT-PCR was extended to serum of patients from low and high infection sites in Zimbabwe and Uganda. The host-derived miRNAs failed to distinguish uninfected from infected individuals. However, analysis of three of the parasite-derived miRNAs (miR-277, miR-3479-3p and bantam) could detect infected individuals from low and high infection intensity sites with specificity/sensitivity values of 89%/80% and 80%/90%, respectively.
Conclusions: This work identifies parasite-derived miRNAs as novel markers of S. mansoni infection in both mice and humans, with the potential to be used with existing techniques to improve S. mansoni diagnosis. In contrast, although host miRNAs are differentially expressed in the liver during infection their abundance levels in serum are variable in human patients and may be useful in cases of extreme pathology but likely hold limited value for detecting prevalence of infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Organisation WH (2013) Schistosomiasis-Fact sheet 115. WHO.
-
- van der Werf MJ, de Vlas SJ, Brooker S, Looman CW, Nagelkerke NJ, et al. (2003) Quantification of clinical morbidity associated with schistosome infection in sub-Saharan Africa. Acta tropica 86: 125–139. - PubMed
-
- King CH, Dickman K, Tisch DJ (2005) Reassessment of the cost of chronic helmintic infection: a meta-analysis of disability-related outcomes in endemic schistosomiasis. Lancet 365: 1561–1569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
