

Brain innate immunity in the regulation of neuroinflammation: therapeutic strategies by modulating CD200-CD200R interaction involve the cannabinoid system
- PMID: 24588829
- PMCID: PMC4157566
- DOI: 10.2174/1381612820666140130202911
Brain innate immunity in the regulation of neuroinflammation: therapeutic strategies by modulating CD200-CD200R interaction involve the cannabinoid system
Abstract
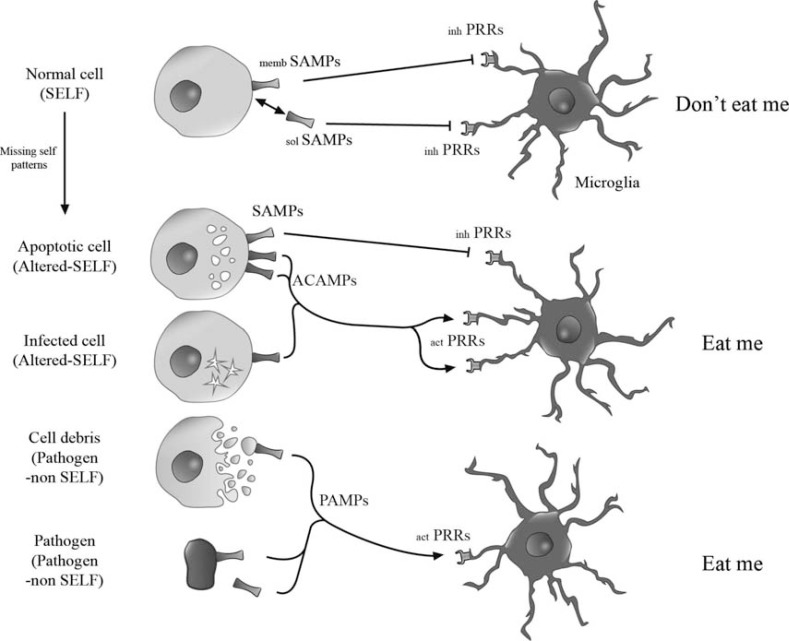
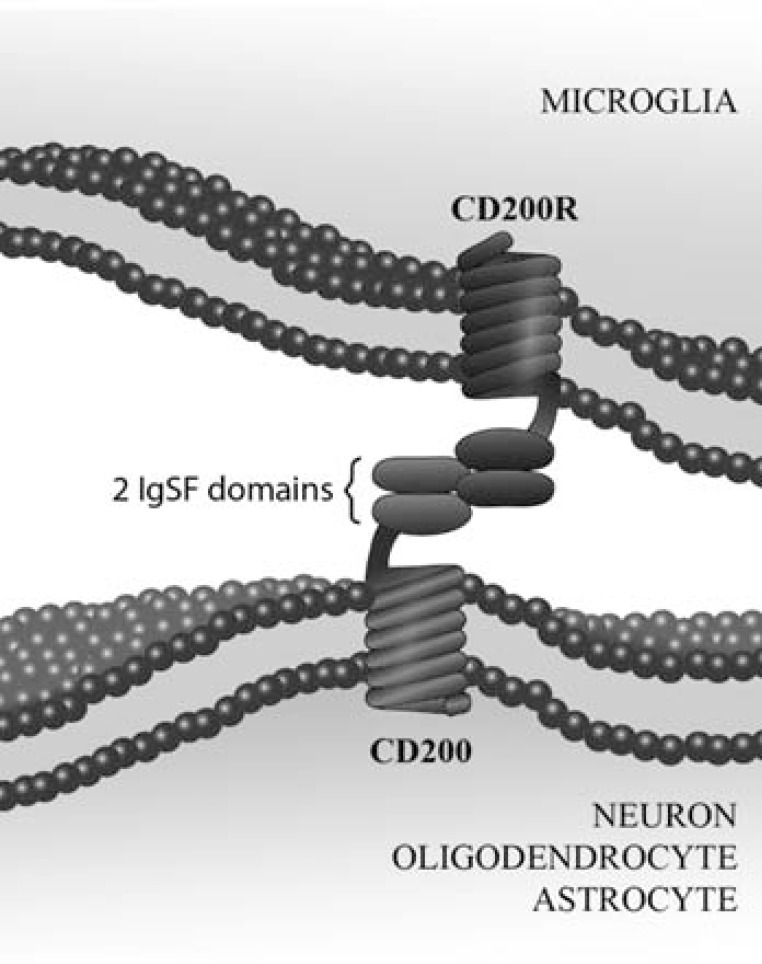
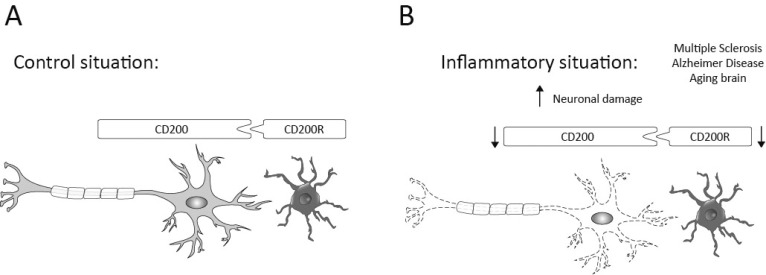
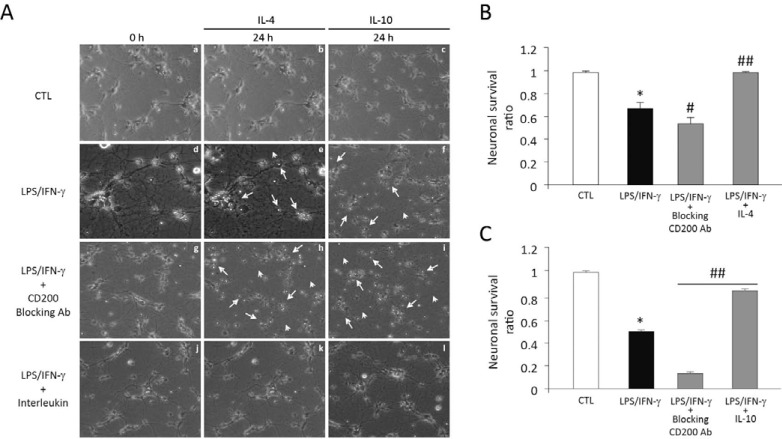
The central nervous system (CNS) innate immune response includes an arsenal of molecules and receptors expressed by professional phagocytes, glial cells and neurons that is involved in host defence and clearance of toxic and dangerous cell debris. However, any uncontrolled innate immune responses within the CNS are widely recognized as playing a major role in the development of autoimmune disorders and neurodegeneration, with multiple sclerosis (MS) Alzheimer's disease (AD) being primary examples. Hence, it is important to identify the key regulatory mechanisms involved in the control of CNS innate immunity and which could be harnessed to explore novel therapeutic avenues. Neuroimmune regulatory proteins (NIReg) such as CD95L, CD200, CD47, sialic acid, complement regulatory proteins (CD55, CD46, fH, C3a), HMGB1, may control the adverse immune responses in health and diseases. In the absence of these regulators, when neurons die by apoptosis, become infected or damaged, microglia and infiltrating immune cells are free to cause injury as well as an adverse inflammatory response in acute and chronic settings. We will herein provide new emphasis on the role of the pair CD200-CD200R in MS and its experimental models: experimental autoimmune encephalomyelitis (EAE) and Theiler's virus induced demyelinating disease (TMEV-IDD). The interest of the cannabinoid system as inhibitor of inflammation prompt us to introduce our findings about the role of endocannabinoids (eCBs) in promoting CD200-CD200 receptor (CD200R) interaction and the benefits caused in TMEV-IDD. Finally, we also review the current data on CD200-CD200R interaction in AD, as well as, in the aging brain.
Figures
Similar articles
-
Activation and control of CNS innate immune responses in health and diseases: a balancing act finely tuned by neuroimmune regulators (NIReg).CNS Neurol Disord Drug Targets. 2011 Feb;10(1):25-43. doi: 10.2174/187152711794488601. CNS Neurol Disord Drug Targets. 2011. PMID: 21143144 Review.
-
CD200-CD200R1 interaction contributes to neuroprotective effects of anandamide on experimentally induced inflammation.Glia. 2012 Sep;60(9):1437-50. doi: 10.1002/glia.22366. Epub 2012 May 31. Glia. 2012. PMID: 22653796
-
Innate immunity and protective neuroinflammation: new emphasis on the role of neuroimmune regulatory proteins.Int Rev Neurobiol. 2007;82:29-55. doi: 10.1016/S0074-7742(07)82002-2. Int Rev Neurobiol. 2007. PMID: 17678954 Review.
-
Distribution of the immune inhibitory molecules CD200 and CD200R in the normal central nervous system and multiple sclerosis lesions suggests neuron-glia and glia-glia interactions.J Neuropathol Exp Neurol. 2009 Feb;68(2):159-67. doi: 10.1097/NEN.0b013e3181964113. J Neuropathol Exp Neurol. 2009. PMID: 19151626
-
Decreased expression of CD200 and CD200 receptor in Alzheimer's disease: a potential mechanism leading to chronic inflammation.Exp Neurol. 2009 Jan;215(1):5-19. doi: 10.1016/j.expneurol.2008.09.003. Epub 2008 Sep 24. Exp Neurol. 2009. PMID: 18938162 Free PMC article.
Cited by
-
Crosstalk between Oxidative Stress and Aging in Neurodegeneration Disorders.Cells. 2023 Feb 27;12(5):753. doi: 10.3390/cells12050753. Cells. 2023. PMID: 36899889 Free PMC article. Review.
-
CD200fc enhances anti-tumoral immune response and inhibits visceral metastasis of breast carcinoma.Oncotarget. 2018 Apr 10;9(27):19147-19158. doi: 10.18632/oncotarget.24931. eCollection 2018 Apr 10. Oncotarget. 2018. PMID: 29721190 Free PMC article.
-
Innovative Therapeutic Strategies for Effective Treatment of Brain Metastases.Int J Mol Sci. 2019 Mar 14;20(6):1280. doi: 10.3390/ijms20061280. Int J Mol Sci. 2019. PMID: 30875730 Free PMC article. Review.
-
Biosafety and mental health: Virus induced cognitive decline.Biosaf Health. 2023 Apr 13;5(3):159-167. doi: 10.1016/j.bsheal.2023.04.002. eCollection 2023 Jun. Biosaf Health. 2023. PMID: 40078510 Free PMC article. Review.
-
Upregulated CD200 in pre-retinal proliferative fibrovascular membranes of proliferative diabetic retinopathy patients and its correlation with vascular endothelial growth factor.Inflamm Res. 2019 Dec;68(12):1071-1079. doi: 10.1007/s00011-019-01290-4. Epub 2019 Oct 14. Inflamm Res. 2019. PMID: 31612255
References
-
- Medzhitov R, Janeway CA., Jr Decoding the patterns of self and nonself by the innate immune system. Science. 2002;296: 298–300. - PubMed
-
- Stuart LM, Stuart R, Ezekowitz A. Phagocytosis elegant complexity. Immunity. 2005;22: 539–50. - PubMed
-
- Nguyen MD, Julien JP, Rivest S. Innate immunity the missing link in neuroprotection and neurodegeneration. Nature Rev Neurosci. 2002;3: 216–27. - PubMed
-
- Elward K, Gasque P. "Eat me" and "don't eat me" signals govern the innate immune response and tissue repair in the CNS emphasis on the critical role of the complement system. Mol Immunol. 2003;40: 85–94. - PubMed
-
- Hauwel M, Furon E, Canova C, Griffiths M, Neal J, Gasque P. Innate (inherent) control of brain infection, brain inflammation and brain repair the role of microglia, astrocytes "protective" glial stem cells, and stromal ependymal cells. Brain Res Rev. 2005;48: 220–33. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
