

Emerging role for nuclear rotation and orientation in cell migration
- PMID: 24589621
- PMCID: PMC3974792
- DOI: 10.4161/cam.27761
Emerging role for nuclear rotation and orientation in cell migration
Abstract
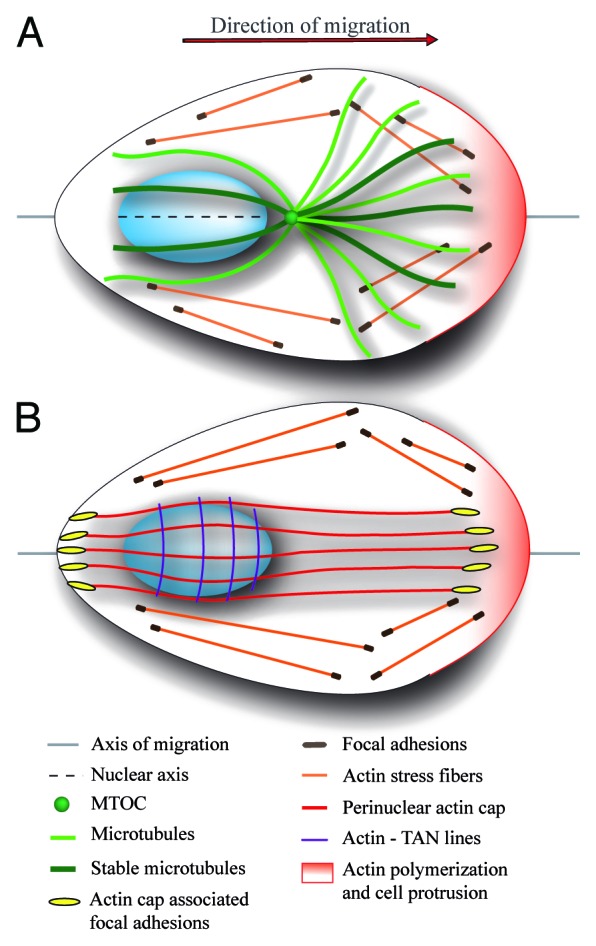
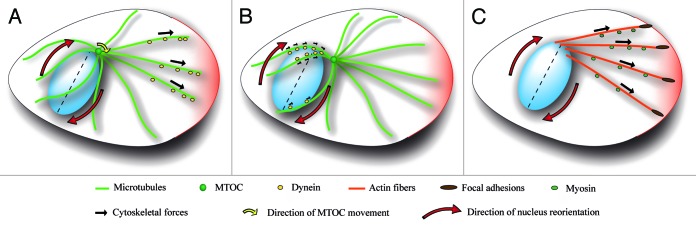
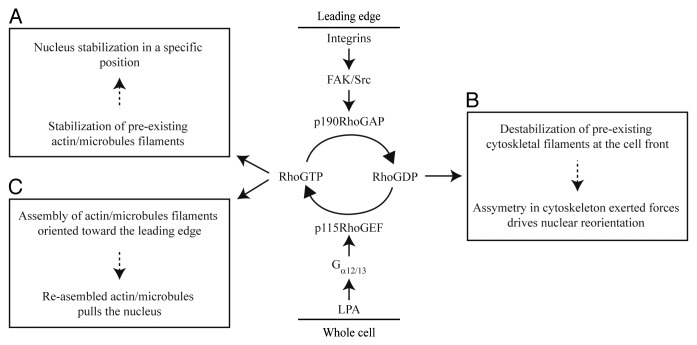
Nucleus movement, positioning, and orientation is precisely specified and actively regulated within cells, and it plays a critical role in many cellular and developmental processes. Mutation of proteins that regulate the nucleus anchoring and movement lead to diverse pathologies, laminopathies in particular, suggesting that the nucleus correct positioning and movement is essential for proper cellular function. In motile cells that polarize toward the direction of migration, the nucleus undergoes controlled rotation promoting the alignment of the nucleus with the axis of migration. Such spatial organization of the cell appears to be optimal for the cell migration. Nuclear reorientation requires the cytoskeleton to be anchored to the nuclear envelope, which exerts pulling or pushing torque on the nucleus. Here we discuss the possible molecular mechanisms regulating the nuclear rotation and reorientation and the significance of this type of nuclear movement for cell migration.
Keywords: FAK; LINC; actin; cell polarity; dynein; focal adhesions; microtubules; migration; myosin; nuclear reorientation.
Figures
Similar articles
-
FAK regulates dynein localisation and cell polarity in migrating mouse fibroblasts.Biol Cell. 2020 Feb;112(2):53-72. doi: 10.1111/boc.201900041. Epub 2020 Jan 9. Biol Cell. 2020. PMID: 31859373
-
The reorientation of cell nucleus promotes the establishment of front-rear polarity in migrating fibroblasts.J Mol Biol. 2013 Jun 12;425(11):2039-2055. doi: 10.1016/j.jmb.2013.02.034. Epub 2013 Mar 21. J Mol Biol. 2013. PMID: 23524135
-
Dorsal stress fibers, transverse actin arcs, and perinuclear actin fibers form an interconnected network that induces nuclear movement in polarizing fibroblasts.FEBS J. 2016 Oct;283(20):3676-3693. doi: 10.1111/febs.13836. Epub 2016 Sep 5. FEBS J. 2016. PMID: 27538255
-
Nuclear movement in fungi.Semin Cell Dev Biol. 2018 Oct;82:3-16. doi: 10.1016/j.semcdb.2017.10.024. Epub 2017 Dec 11. Semin Cell Dev Biol. 2018. PMID: 29241689 Free PMC article. Review.
-
Connecting the nucleus to the cytoskeleton for nuclear positioning and cell migration.Adv Exp Med Biol. 2014;773:505-20. doi: 10.1007/978-1-4899-8032-8_23. Adv Exp Med Biol. 2014. PMID: 24563363 Review.
Cited by
-
Altered mechanotransduction in adolescent idiopathic scoliosis osteoblasts: an exploratory in vitro study.Sci Rep. 2022 Feb 3;12(1):1846. doi: 10.1038/s41598-022-05918-0. Sci Rep. 2022. PMID: 35115632 Free PMC article.
-
Curvature Perception of Mesenchymal Cells on Mesoscale Topographies.Adv Healthc Mater. 2025 Jan;14(3):e2402865. doi: 10.1002/adhm.202402865. Epub 2024 Dec 10. Adv Healthc Mater. 2025. PMID: 39659136 Free PMC article.
-
Physical Forces and Transient Nuclear Envelope Rupture during Metastasis: The Key for Success?Cancers (Basel). 2021 Dec 24;14(1):83. doi: 10.3390/cancers14010083. Cancers (Basel). 2021. PMID: 35008251 Free PMC article. Review.
-
Chitosan Micro-Grooved Membranes with Increased Asymmetry for the Improvement of the Schwann Cell Response in Nerve Regeneration.Int J Mol Sci. 2021 Jul 23;22(15):7901. doi: 10.3390/ijms22157901. Int J Mol Sci. 2021. PMID: 34360664 Free PMC article.
-
The Interplay Between Cell-Cell and Cell-Matrix Forces Regulates Cell Migration Dynamics.Biophys J. 2019 Nov 19;117(10):1795-1804. doi: 10.1016/j.bpj.2019.10.015. Epub 2019 Oct 23. Biophys J. 2019. PMID: 31706566 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous