

A novel lily anther-specific gene encodes adhesin-like proteins associated with exine formation during anther development
- PMID: 24591055
- PMCID: PMC3991738
- DOI: 10.1093/jxb/eru051
A novel lily anther-specific gene encodes adhesin-like proteins associated with exine formation during anther development
Abstract
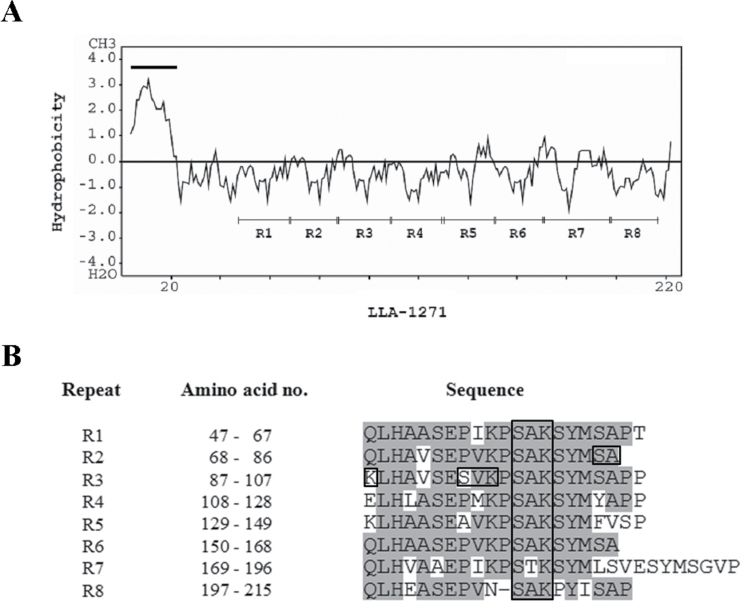
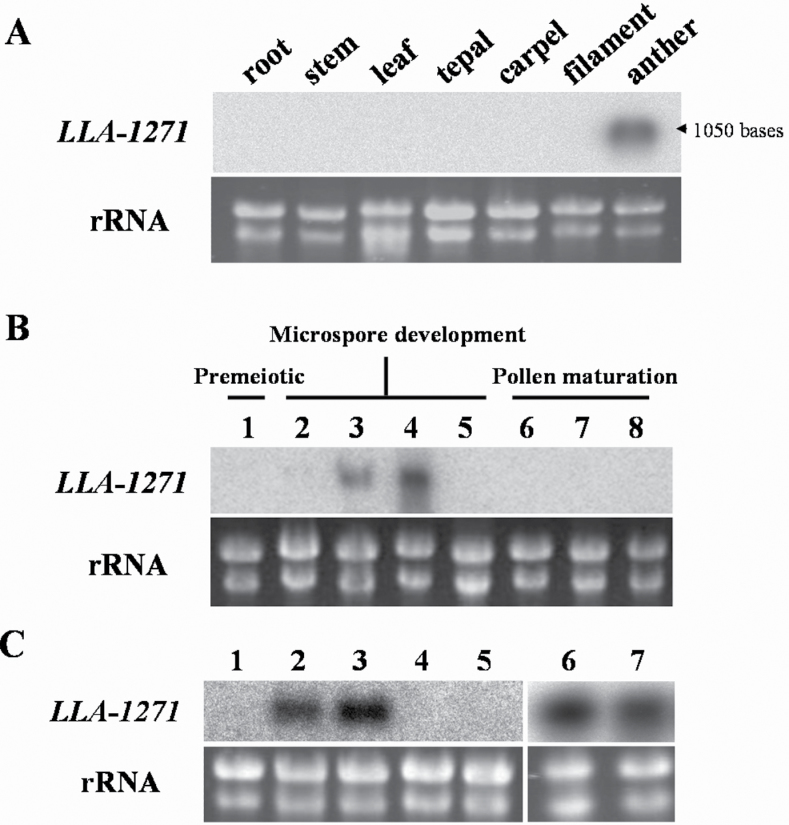
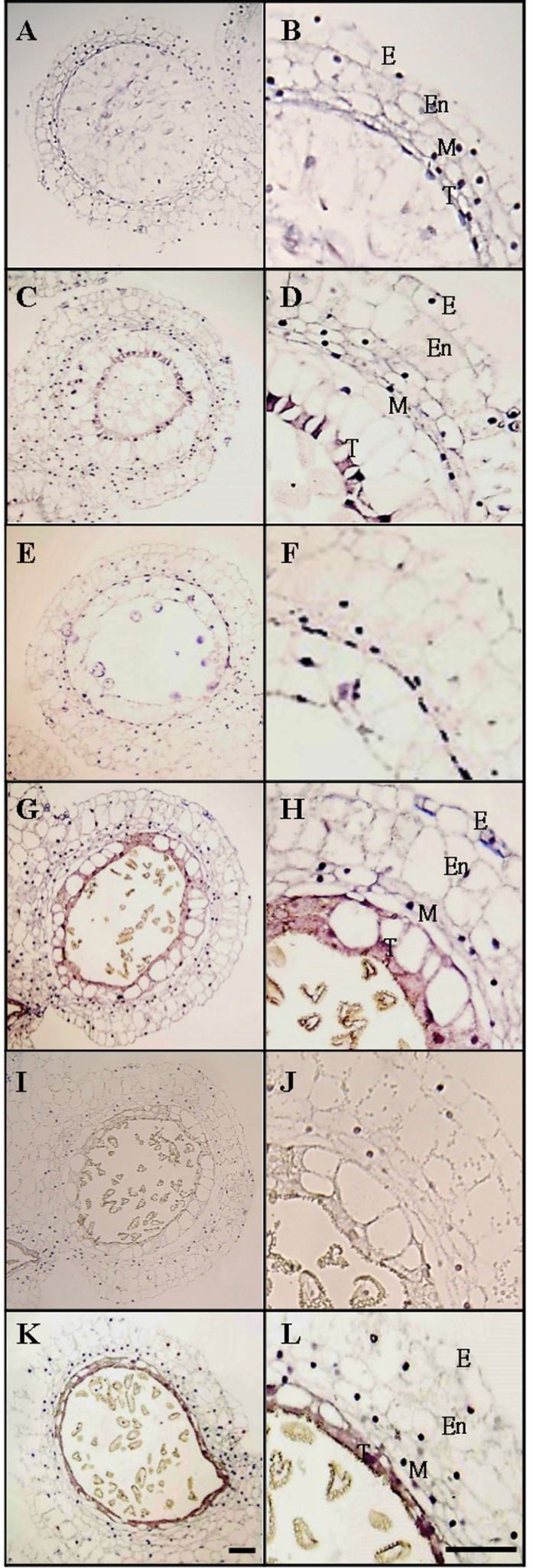
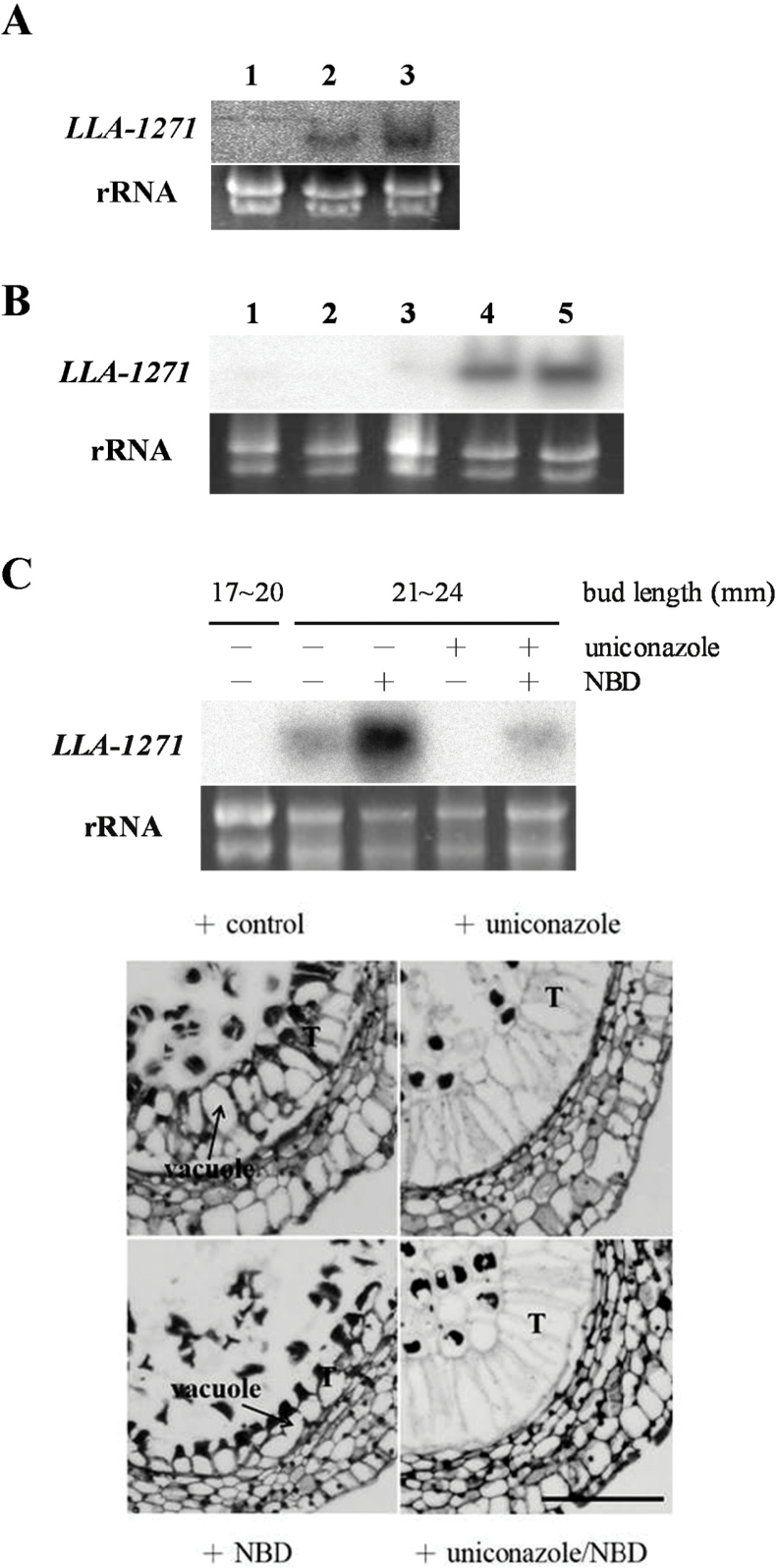
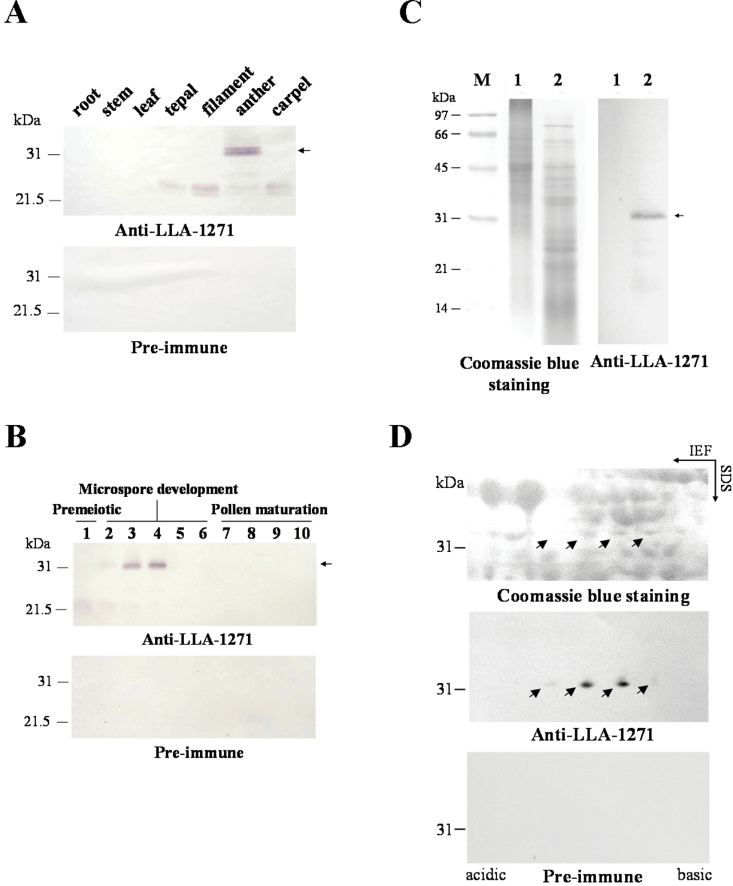
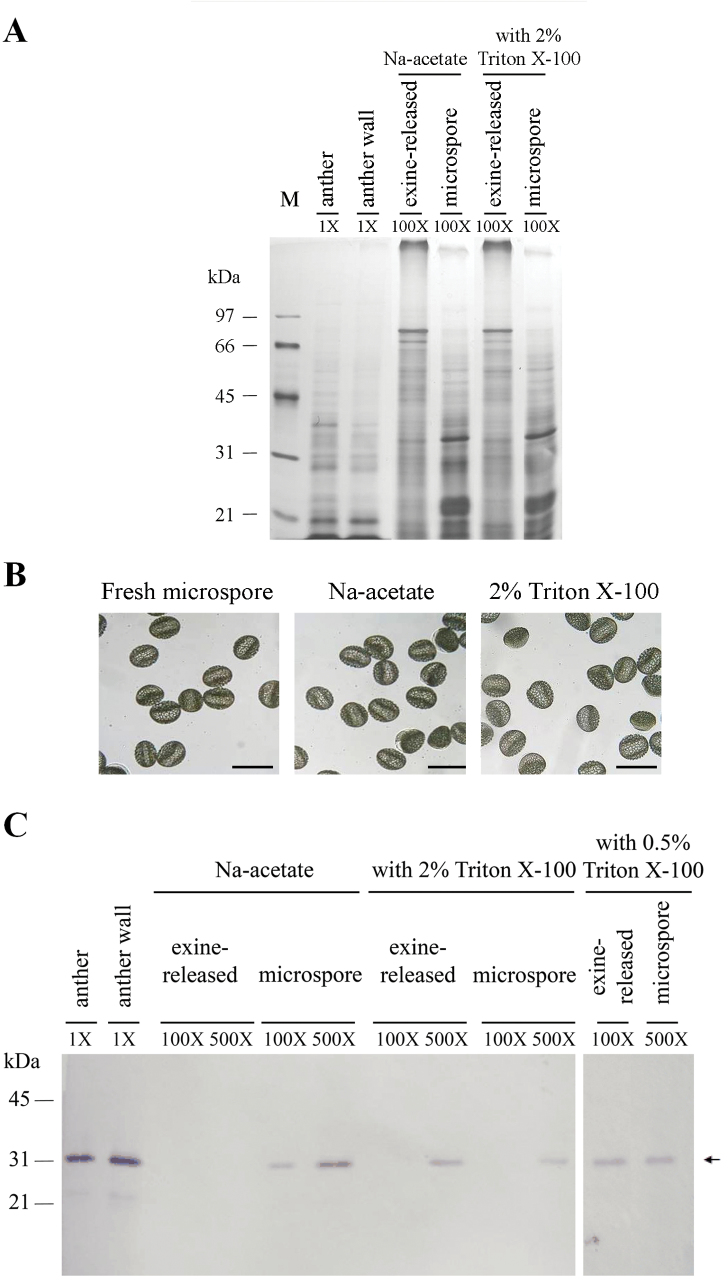
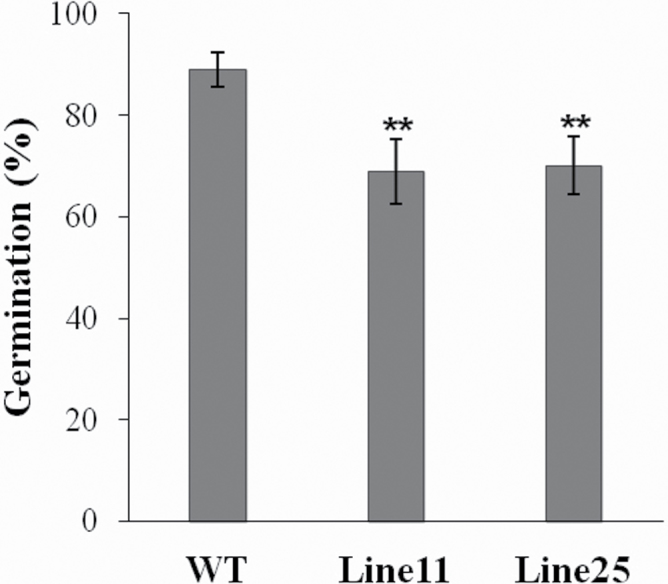
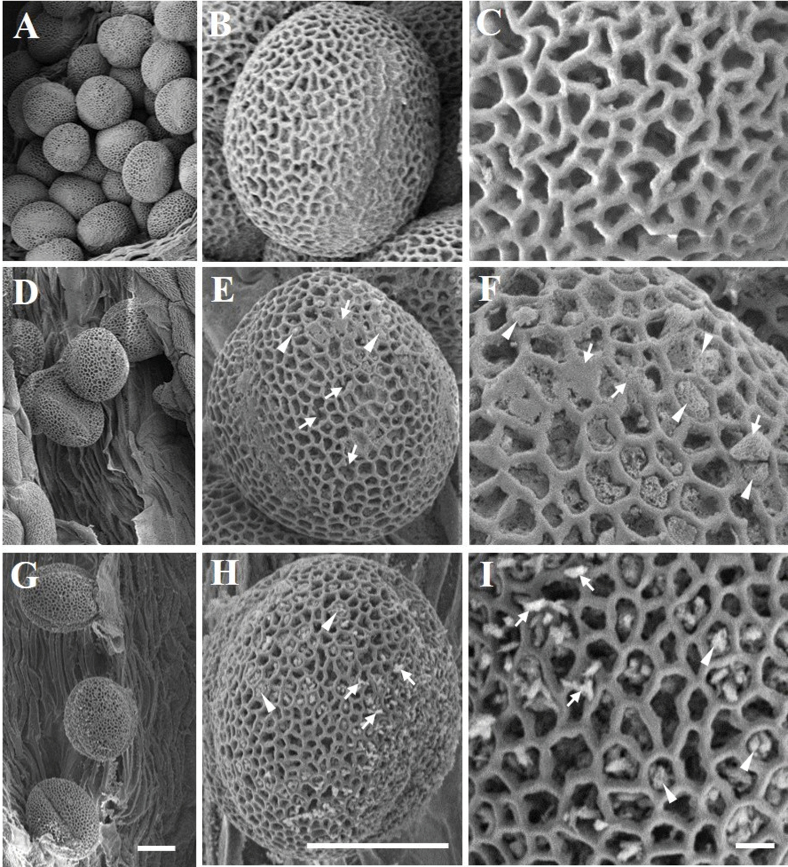
The anther-specific gene LLA1271 isolated from lily (Lilium longiflorum Thunb.) anthers is novel and exists in two forms. The protein encoded by LLA1271 may represent an adhesin-like protein first found in higher plants. The protein contains a typical N-terminal signal peptide followed by a highly conserved repeat domain. The LLA1271 gene is temporally expressed at the phase of microspore development. RNA blot and RNA in situ hybridization analyses demonstrated that the gene was expressed both in the tapetum and in the microspore. The gene is endo- and exogenously induced by gibberellin. Studies with the gibberellin biosynthesis inhibitor uniconazole and an inhibitor of ethylene activity, 2,5-norbornadien (NBD), revealed that LLA1271 is negatively regulated by ethylene, and a cross-talk of regulation between gibberellin and ethylene occurs in young anthers. The treatment with NBD caused the tapetum to become densely cytoplasmic and highly polarized, whereas uniconazole arrested tapetal development in a state close to that of a tapetum without treatment. The LLA1271 protein is heat stable and heterogeneous. An immunoblot of separated protein fractions of the anther revealed that the LLA1271 protein was detected in protein fraction of the microspore released from the cell wall by treatment with either 0.5% or 2% Triton X-100. Ectopic expression of LLA1271 resulted in impaired stamen and low pollen germination. Scanning electron microscopy of TAP::LLA1271 pollen showed distorted exine formation and patterning. The LLA1271 protein once synthesized in both the tapetum and microspore is secreted and deposited on the surface of microspores, moderately affecting exine formation and patterning.
Keywords: Adhesin; anther; exine; hormone; lily (Lilium longiflorum Thunb.); microspore; tapetum..
Figures


References
-
- Alexander MP. 1969. Differential staining of aborted and nonaborted pollen. Stain Technology 44, 117–122 - PubMed
-
- Aouali N, Laporte P, Clément C. 2001. Pectin secretion and distribution in the anther during pollen development in Lilium. Planta 213, 71–79 - PubMed
-
- Atanassov I, Russinova E, Antonov L, Atanassov A. 1998. Expression of an anther-specific chalcone synthase-like gene is correlated with uninucleate microspore development in Nicotiana sylvestris . Plant Molecular Biology 38, 1169–1178 - PubMed
-
- Bechtold N, Ellis J, Pelletier G. 1993. In planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. Comptes Rendus de l’Académie des Sciences Series III 316, 1194–1199
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
