

Identification of NuRSERY, a new functional HDAC complex composed by HDAC5, GATA1, EKLF and pERK present in human erythroid cells
- PMID: 24594363
- PMCID: PMC4003889
- DOI: 10.1016/j.biocel.2014.02.019
Identification of NuRSERY, a new functional HDAC complex composed by HDAC5, GATA1, EKLF and pERK present in human erythroid cells
Abstract
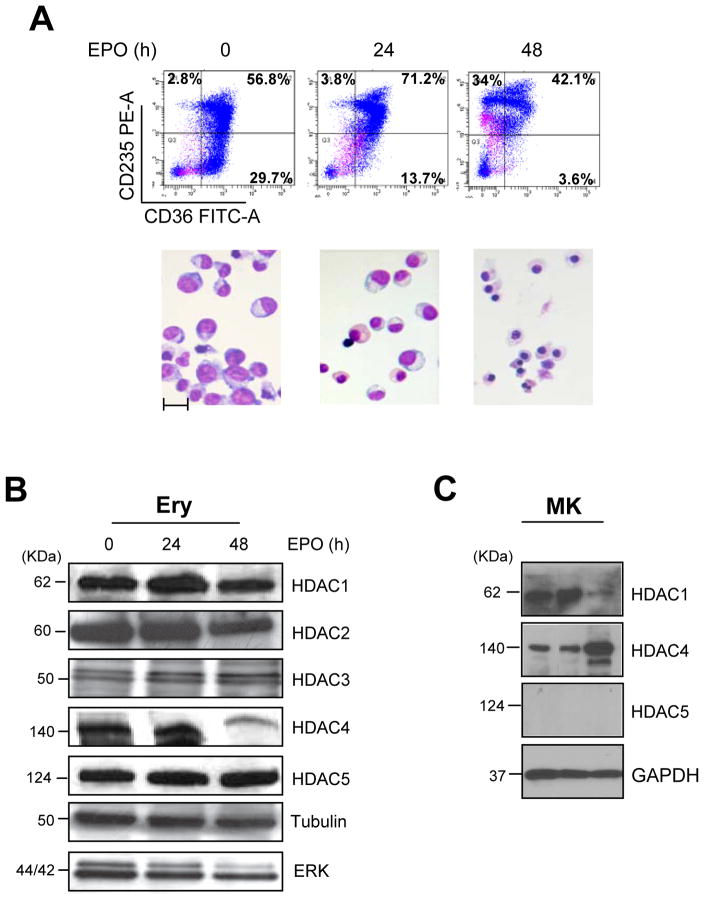
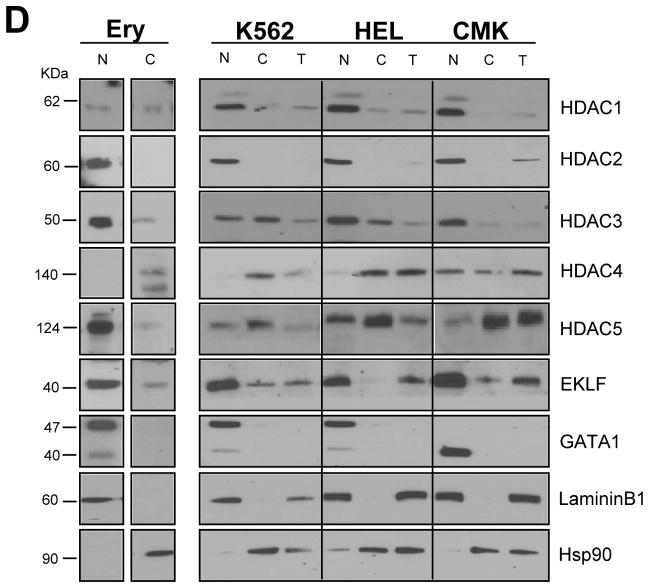
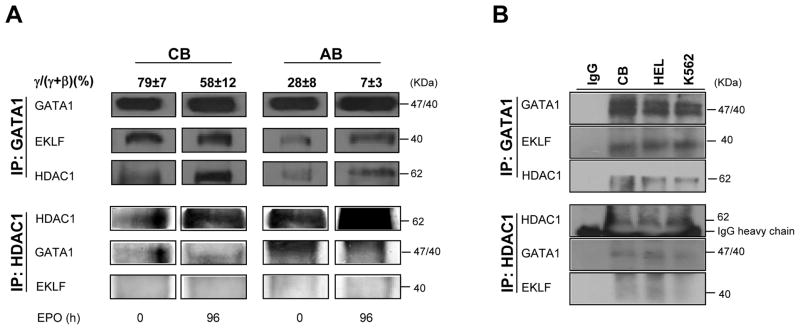
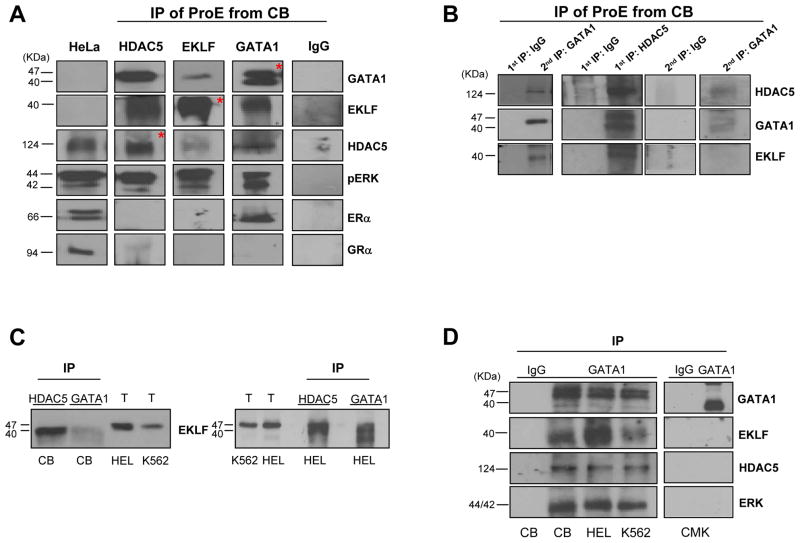
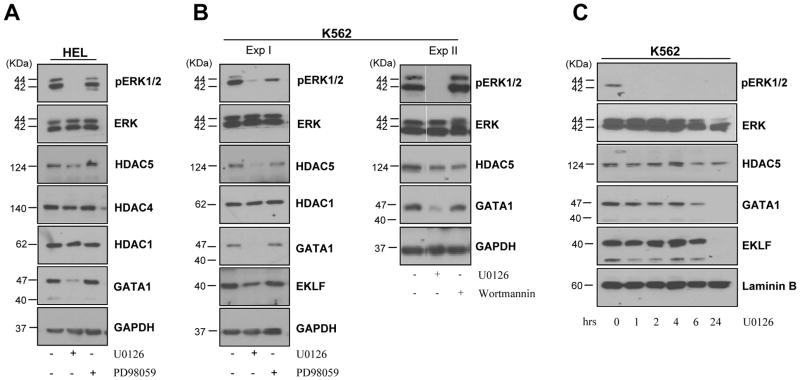
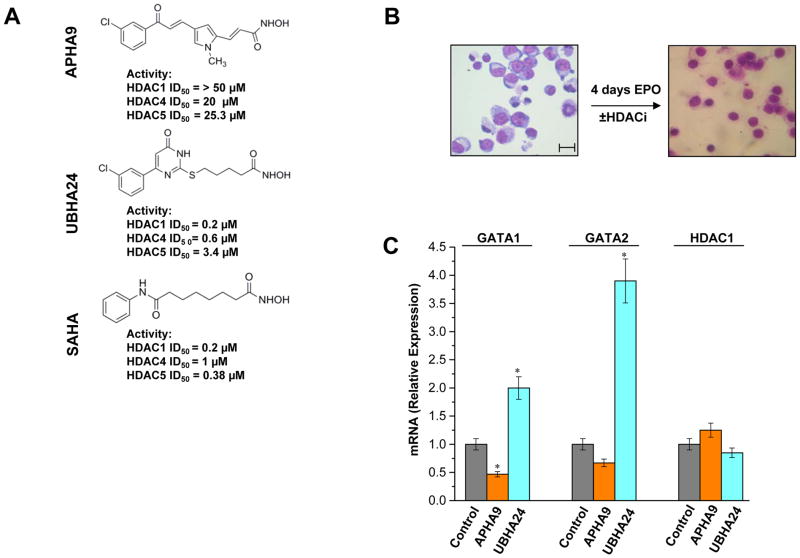
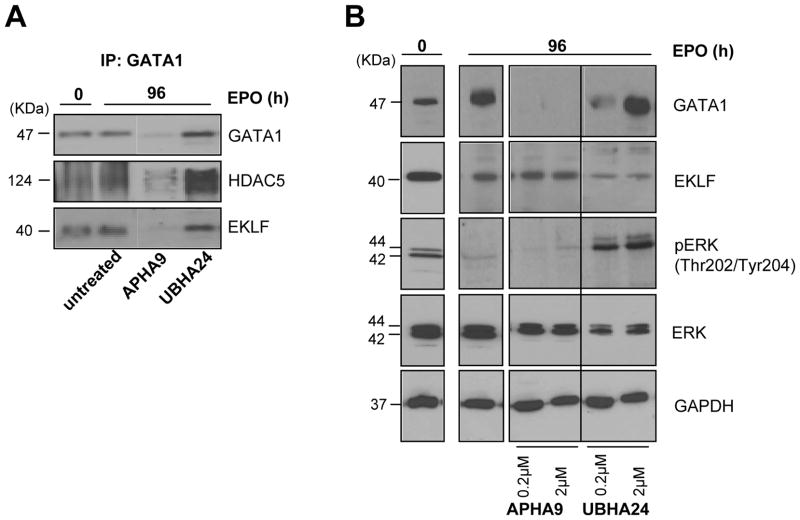
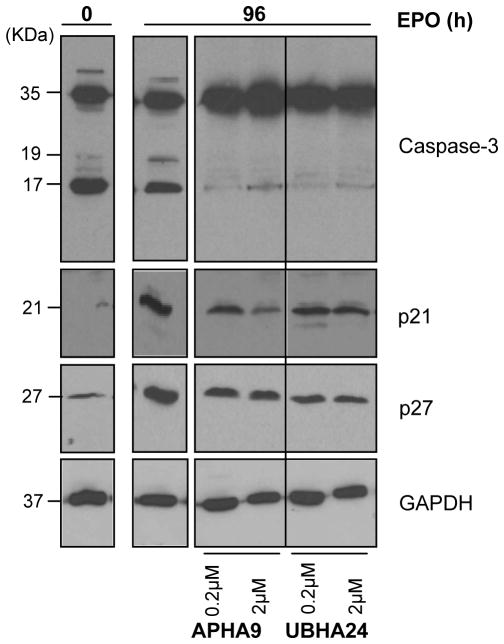
To clarify the role of HDACs in erythropoiesis, expression, activity and function of class I (HDAC1, HDAC2, HDAC3) and class IIa (HDAC4, HDAC5) HDACs during in vitro maturation of human erythroblasts were compared. During erythroid maturation, expression of HDAC1, HDAC2 and HDAC3 remained constant and activity and GATA1 association (its partner of the NuRD complex), of HDAC1 increased. By contrast, HDAC4 content drastically decreased and HDAC5 remained constant in content but decreased in activity. In erythroid cells, pull down experiments identified the presence of a novel complex formed by HDAC5, GATA1, EKLF and pERK which was instead undetectable in cells of the megakaryocytic lineage. With erythroid maturation, association among HDAC5, GATA1 and EKLF persisted but levels of pERK sharply decreased. Treatment of erythroleukemic cells with inhibitors of ERK phosphorylation reduced by >90% the total and nuclear content of HDAC5, GATA1 and EKLF, suggesting that ERK phosphorylation is required for the formation of this complex. Based on the function of class IIa HDACs as chaperones of other proteins to the nucleus and the erythroid-specificity of HDAC5 localization, this novel HDAC complex was named nuclear remodeling shuttle erythroid (NuRSERY). Exposure of erythroid cells to the class II-selective HDAC inhibitor (HDACi) APHA9 increased γ/(γ+β) globin expression ratios (Mai et al., 2007), suggesting that NuRSERY may regulate globin gene expression. In agreement with this hypothesis, exposure of erythroid cells to APHA9 greatly reduced the association among HDAC5, GATA1 and EKLF. Since exposure to APHA9 did not affect survival rates or p21 activation, NuRSERY may represent a novel, possibly less toxic, target for epigenetic therapies of hemoglobinopaties and other disorders.
Keywords: EKLF; Erythropoiesis; GATA1; Histone deacetylase inhibitors; Histone deacetylases.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declared no competing financial or other interests.
The authors have declared that no competing interests exists
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
