

ZapE is a novel cell division protein interacting with FtsZ and modulating the Z-ring dynamics
- PMID: 24595368
- PMCID: PMC3958796
- DOI: 10.1128/mBio.00022-14
ZapE is a novel cell division protein interacting with FtsZ and modulating the Z-ring dynamics
Abstract
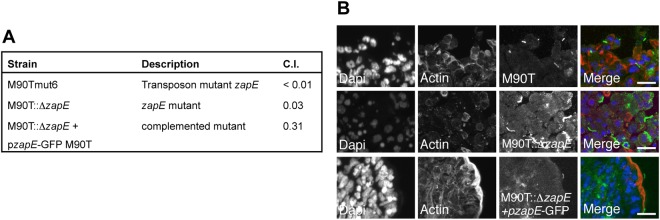
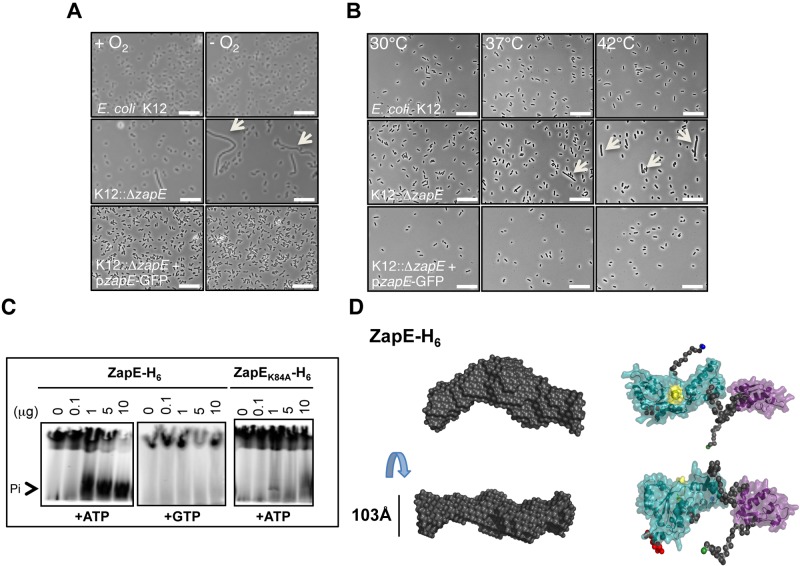
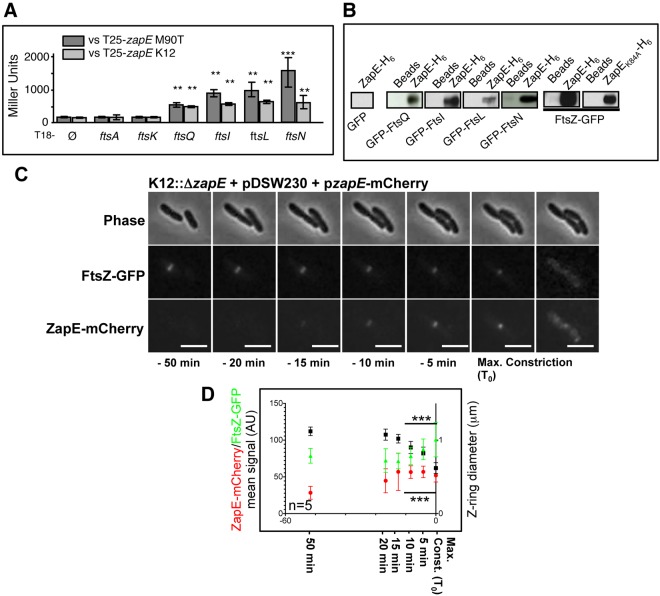
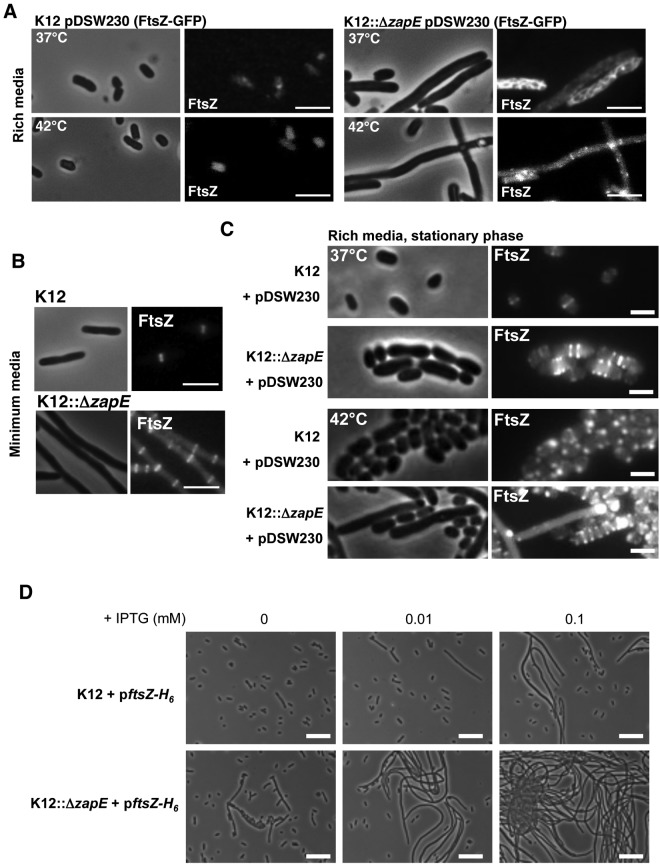
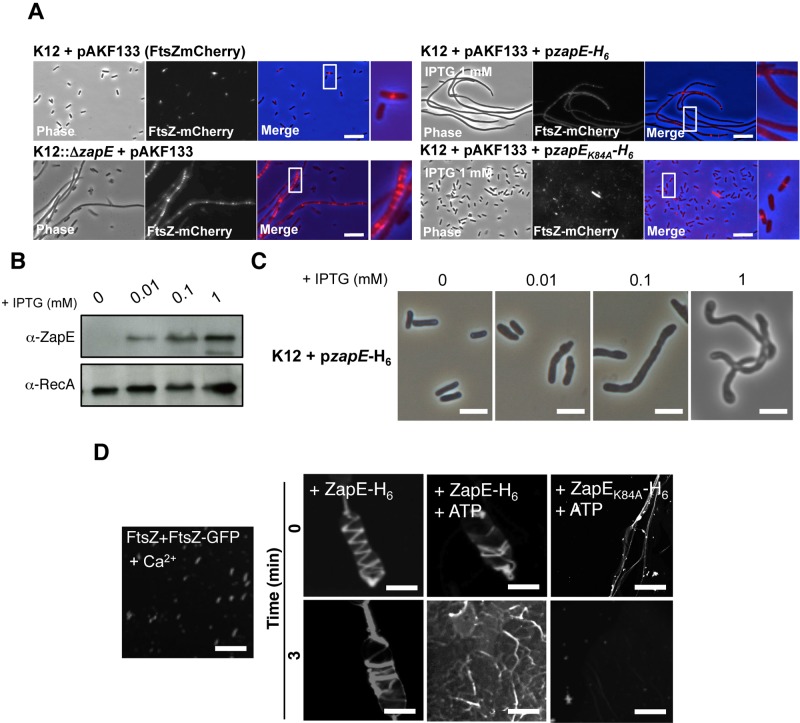
Bacterial cell division requires the formation of a mature divisome complex positioned at the midcell. The localization of the divisome complex is determined by the correct positioning, assembly, and constriction of the FtsZ ring (Z-ring). Z-ring constriction control remains poorly understood and (to some extent) controversial, probably due to the fact that this phenomenon is transient and controlled by numerous factors. Here, we characterize ZapE, a novel ATPase found in Gram-negative bacteria, which is required for growth under conditions of low oxygen, while loss of zapE results in temperature-dependent elongation of cell shape. We found that ZapE is recruited to the Z-ring during late stages of the cell division process and correlates with constriction of the Z-ring. Overexpression or inactivation of zapE leads to elongation of Escherichia coli and affects the dynamics of the Z-ring during division. In vitro, ZapE destabilizes FtsZ polymers in an ATP-dependent manner. IMPORTANCE Bacterial cell division has mainly been characterized in vitro. In this report, we could identify ZapE as a novel cell division protein which is not essential in vitro but is required during an infectious process. The bacterial cell division process relies on the assembly, positioning, and constriction of FtsZ ring (the so-called Z-ring). Among nonessential cell division proteins recently identified, ZapE is the first in which detection at the Z-ring correlates with its constriction. We demonstrate that ZapE abundance has to be tightly regulated to allow cell division to occur; absence or overexpression of ZapE leads to bacterial filamentation. As zapE is not essential, we speculate that additional Z-ring destabilizing proteins transiently recruited during late cell division process might be identified in the future.
Figures
Similar articles
-
Structure and Mutational Analyses of Escherichia coli ZapD Reveal Charged Residues Involved in FtsZ Filament Bundling.J Bacteriol. 2016 May 13;198(11):1683-1693. doi: 10.1128/JB.00969-15. Print 2016 Jun 1. J Bacteriol. 2016. PMID: 27021560 Free PMC article.
-
MinC N- and C-Domain Interactions Modulate FtsZ Assembly, Division Site Selection, and MinD-Dependent Oscillation in Escherichia coli.J Bacteriol. 2019 Jan 28;201(4):e00374-18. doi: 10.1128/JB.00374-18. Print 2019 Feb 15. J Bacteriol. 2019. PMID: 30455283 Free PMC article.
-
Cell Cycle-Dependent Recruitment of FtsN to the Divisome in Escherichia coli.mBio. 2022 Aug 30;13(4):e0201722. doi: 10.1128/mbio.02017-22. Epub 2022 Aug 15. mBio. 2022. PMID: 35968943 Free PMC article.
-
Assembly dynamics of the bacterial MinCDE system and spatial regulation of the Z ring.Annu Rev Biochem. 2007;76:539-62. doi: 10.1146/annurev.biochem.75.103004.142652. Annu Rev Biochem. 2007. PMID: 17328675 Review.
-
Z ring as executor of bacterial cell division.J Mol Microbiol Biotechnol. 2006;11(3-5):140-51. doi: 10.1159/000094050. J Mol Microbiol Biotechnol. 2006. PMID: 16983191 Review.
Cited by
-
Melatonin Treatment Inhibits the Growth of Xanthomonas oryzae pv. oryzae.Front Microbiol. 2018 Oct 4;9:2280. doi: 10.3389/fmicb.2018.02280. eCollection 2018. Front Microbiol. 2018. PMID: 30337911 Free PMC article.
-
Throwing a spotlight on genomic dark matter: The power and potential of transposon-insertion sequencing.J Biol Chem. 2025 Jun;301(6):110231. doi: 10.1016/j.jbc.2025.110231. Epub 2025 May 14. J Biol Chem. 2025. PMID: 40378959 Free PMC article. Review.
-
ZapE/Afg1 interacts with Oxa1 and its depletion causes a multifaceted phenotype.PLoS One. 2020 Jun 24;15(6):e0234918. doi: 10.1371/journal.pone.0234918. eCollection 2020. PLoS One. 2020. PMID: 32579605 Free PMC article.
-
The Hfq regulon of Neisseria meningitidis.FEBS Open Bio. 2017 Apr 25;7(6):777-788. doi: 10.1002/2211-5463.12218. eCollection 2017 Jun. FEBS Open Bio. 2017. PMID: 28593133 Free PMC article.
-
Beyond the Discovery Void: New targets for antibacterial compounds.Sci Prog. 2016 Jun 1;99(2):153-182. doi: 10.3184/003685016X14616130512308. Sci Prog. 2016. PMID: 28742471 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
