

OsmiR396d-regulated OsGRFs function in floral organogenesis in rice through binding to their targets OsJMJ706 and OsCR4
- PMID: 24596329
- PMCID: PMC4012577
- DOI: 10.1104/pp.114.235564
OsmiR396d-regulated OsGRFs function in floral organogenesis in rice through binding to their targets OsJMJ706 and OsCR4
Abstract
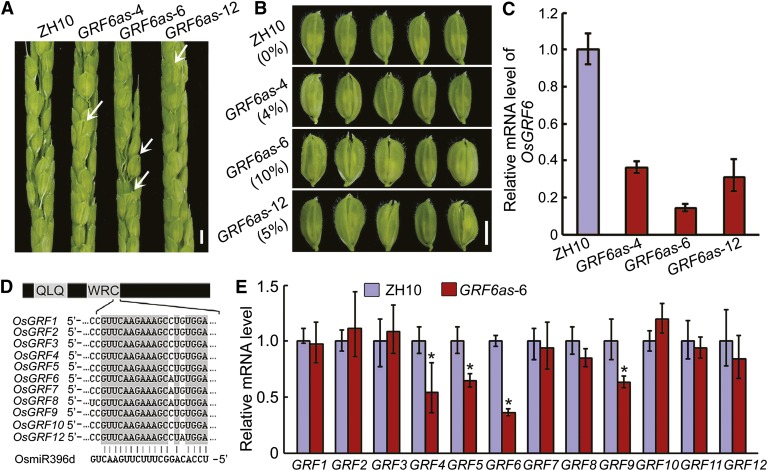
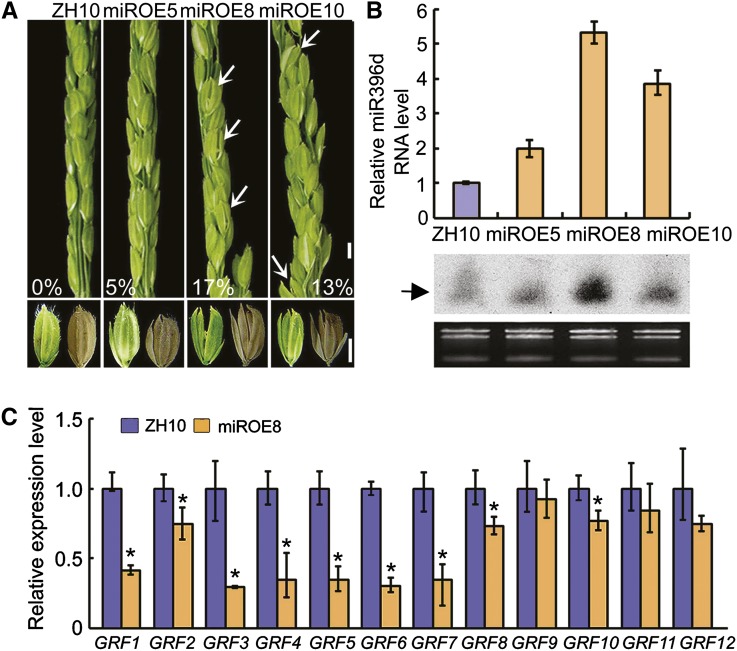
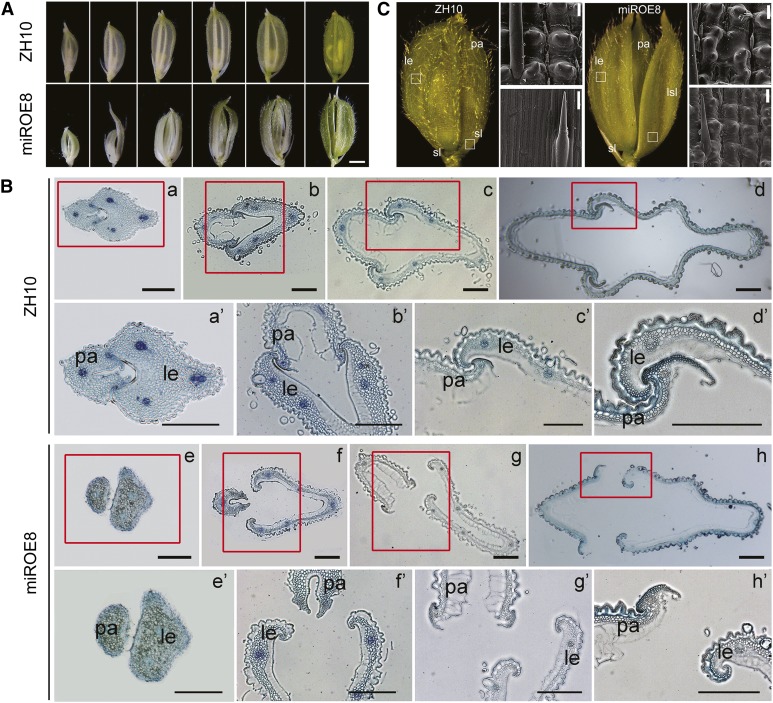
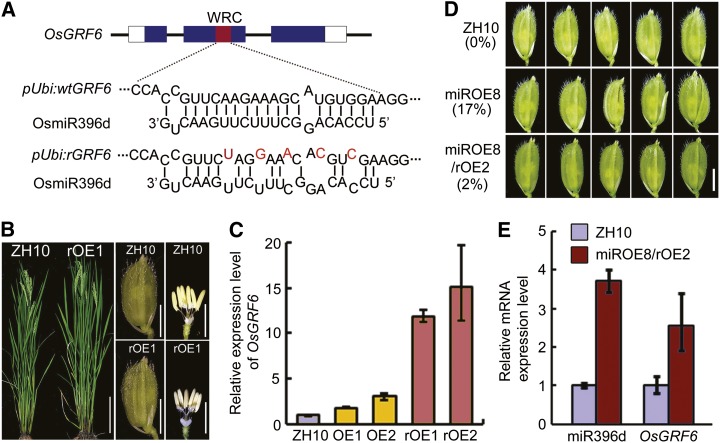
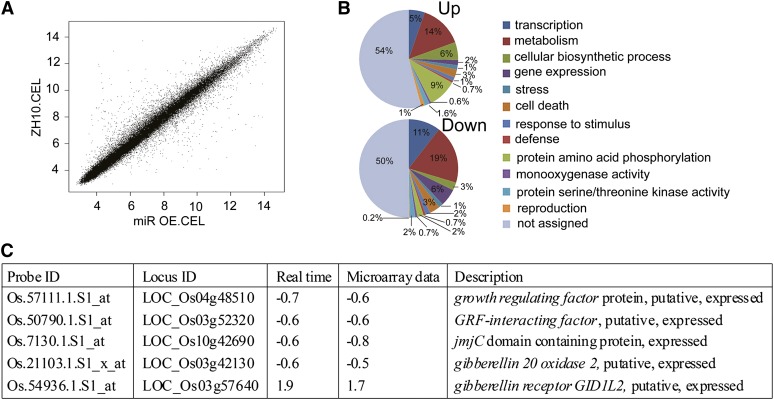
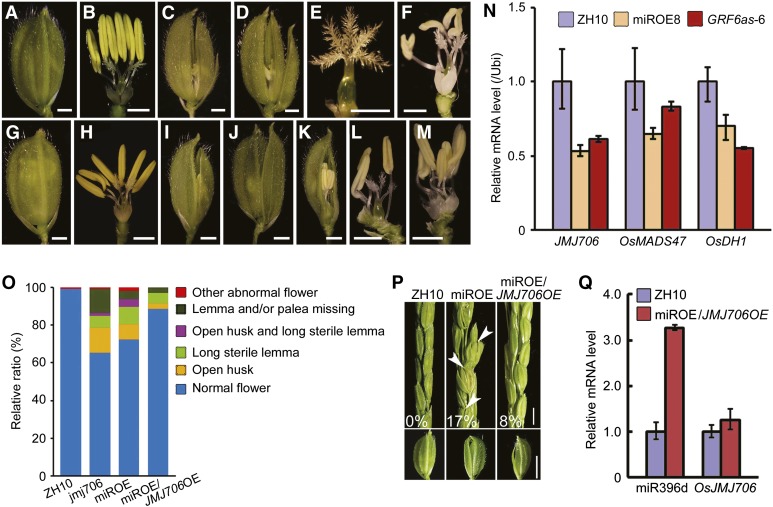
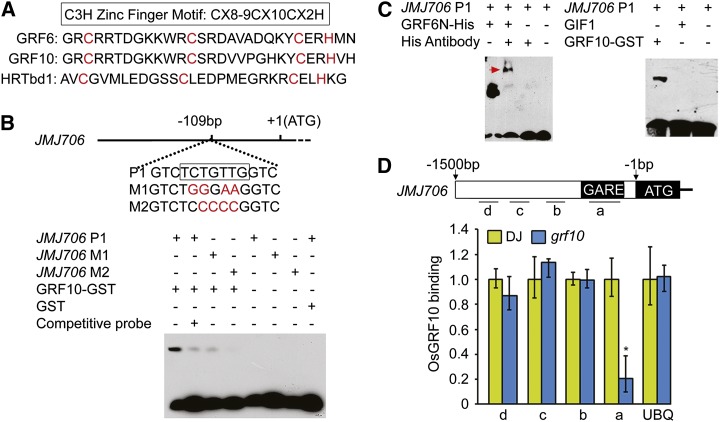
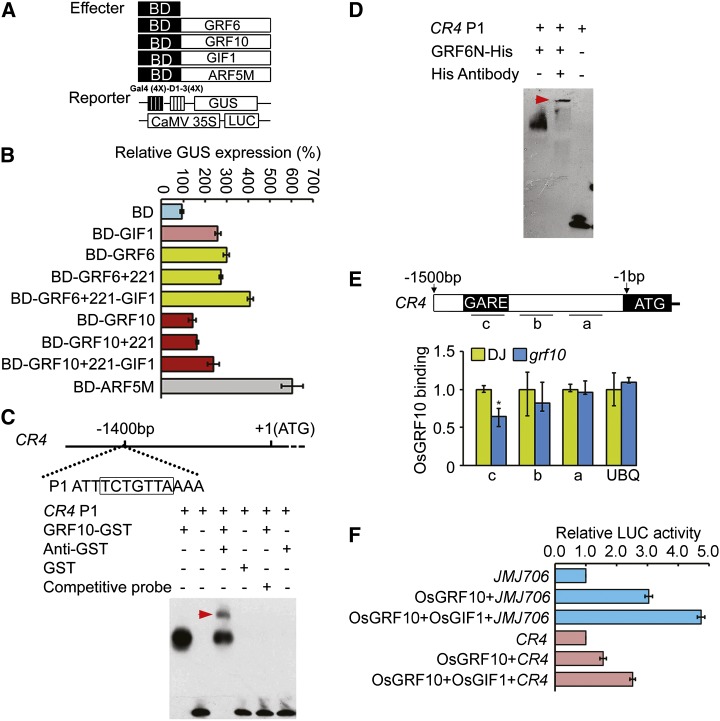
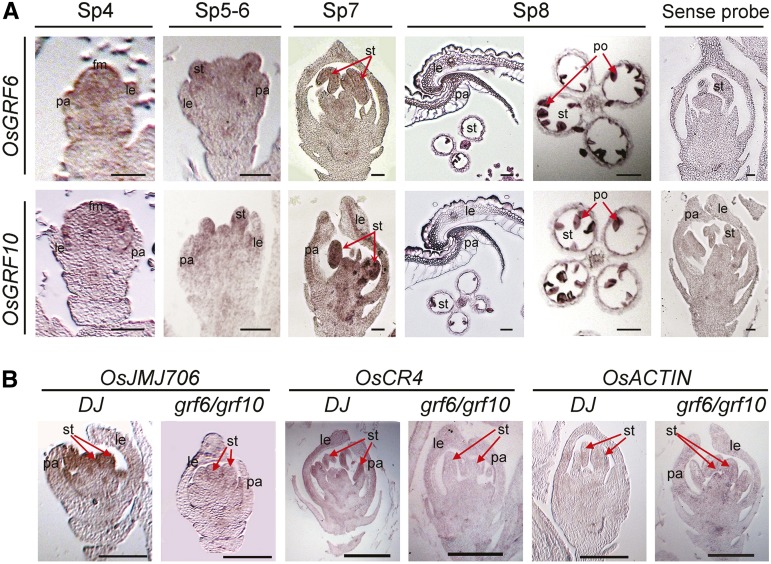
Inflorescence and spikelet development determine grain yields in cereals. Although multiple genes are known to be involved in the regulation of floral organogenesis, the underlying molecular network remains unclear in cereals. Here, we report that the rice (Oryza sativa) microRNA396d (OsmiR396d) and its Os Growth Regulating Factor (OsGRF) targets, together with Os Growth Regulating Factor-Interacting Factor1 (OsGIF1), are involved in the regulation of floral organ development through the rice JMJD2 family jmjC gene 706 (OsJMJ706) and crinkly4 receptor-like kinase (OsCR4). Transgenic knockdown lines of OsGRF6, a predicted target of OsmiR396d, and overexpression lines of OsmiR396d showed similar defects in floral organ development, including open husks, long sterile lemmas, and altered floral organ morphology. These defects were almost completely rescued by overexpression of OsGRF6. OsGRF6 and its ortholog OsGRF10 were the most highly expressed OsGRF family members in young inflorescences, and the grf6/grf10 double mutant displayed abnormal florets. OsGRF6/OsGRF10 localized to the nucleus, and electrophoretic mobility shift assays revealed that both OsGRF6 and OsGRF10 bind the GA response element in the promoters of OsJMJ706 and OsCR4, which were reported to participate in the regulation of floral organ development. In addition, OsGRF6 and OsGRF10 could transactivate OsJMJ706 and OsCR4, an activity that was enhanced in the presence of OsGIF1, which can bind both OsGRF6 and OsGRF10. Together, our results suggest that OsmiR396d regulates the expression of OsGRF genes, which function with OsGIF1 in floret development through targeting of JMJ706 and OsCR4. This work thus reveals a microRNA-mediated regulation module for controlling spikelet development in rice.
Figures


References
-
- Bommert P, Satoh-Nagasawa N, Jackson D, Hirano HY. (2005) Genetics and evolution of inflorescence and flower development in grasses. Plant Cell Physiol 46: 69–78 - PubMed
-
- Choi D, Kim JH, Kende H. (2004) Whole genome analysis of the OsGRF gene family encoding plant-specific putative transcription activators in rice (Oryza sativa L.). Plant Cell Physiol 45: 897–904 - PubMed
-
- Cui R, Han J, Zhao S, Su K, Wu F, Du X, Xu Q, Chong K, Theissen G, Meng Z. (2010) Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). Plant J 61: 767–781 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
