

Do fish oil omega-3 fatty acids enhance antioxidant capacity and mitochondrial fatty acid oxidation in human atrial myocardium via PPARγ activation?
- PMID: 24597798
- PMCID: PMC4142835
- DOI: 10.1089/ars.2014.5888
Do fish oil omega-3 fatty acids enhance antioxidant capacity and mitochondrial fatty acid oxidation in human atrial myocardium via PPARγ activation?
Abstract
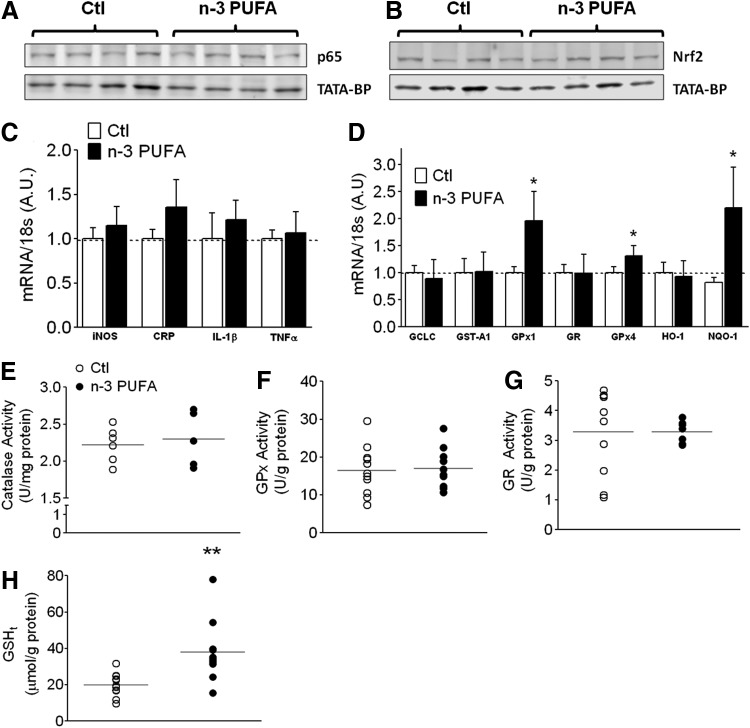
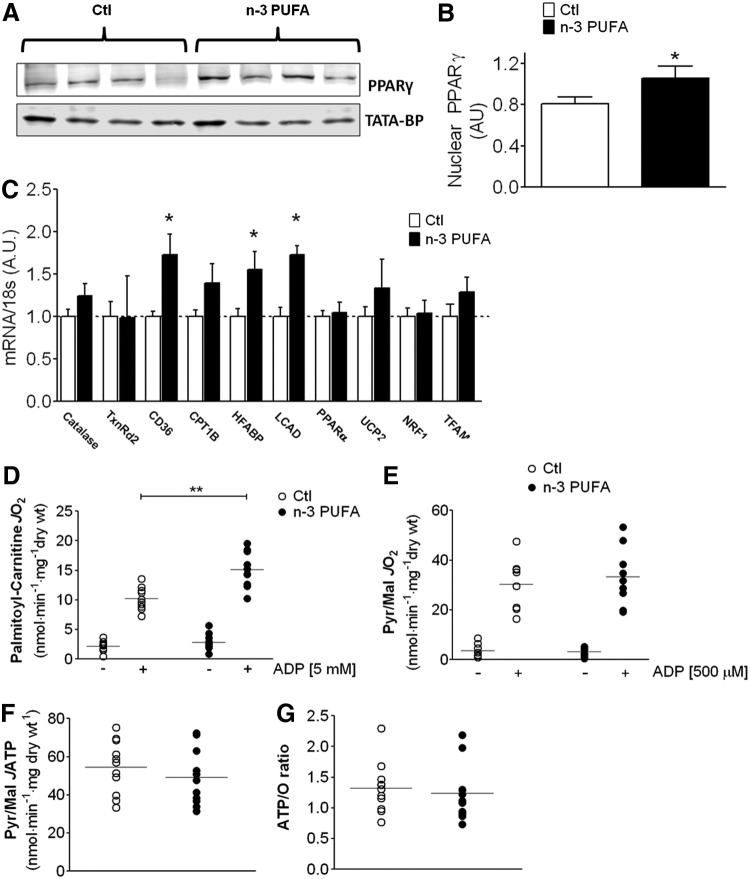
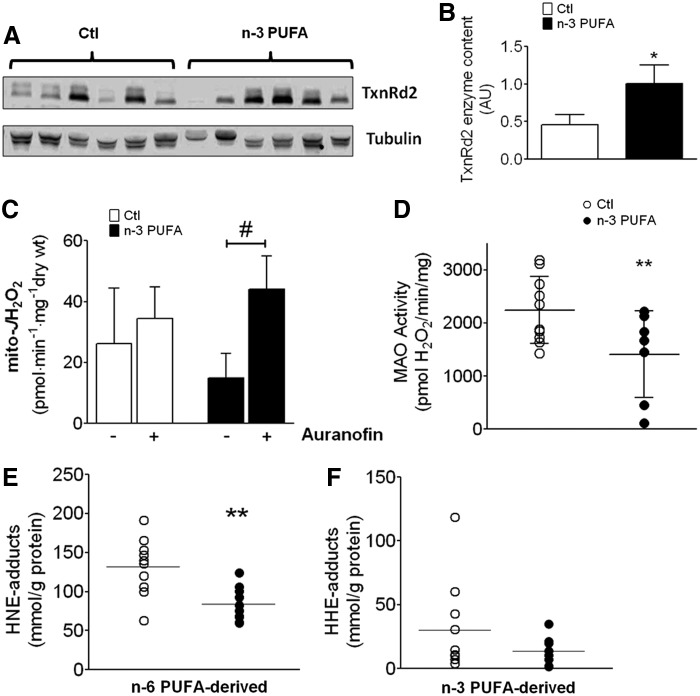
Abstract Studies in experimental models suggest that n-3 polyunsaturated fatty acids (PUFAs) improve metabolic and anti-inflammatory/antioxidant capacity of the heart, although the mechanisms are unclear and translational evidence is lacking. In this study, patients ingested a moderately high dose of n-3 PUFAs (3.4 g/day eicosapentaenoic (EPA) and doxosahexaenoic acid (DHA) ethyl-esters) for a period of 2-3 weeks before having elective cardiac surgery. Blood was obtained before treatment and at the time of surgery, and myocardial tissue from the right atrium was also dissected during surgery. Blood EPA levels increased and myocardial tissue EPA and DHA levels were significantly higher in n-3 PUFA-treated patients compared with untreated, standard-of-care control patients. Interestingly, n-3 PUFA patients had greater nuclear transactivation of peroxisome proliferator-activated receptor-γ (PPARγ), fatty acid metabolic gene expression, and enhanced mitochondrial respiration supported by palmitoyl-carnitine in the atrial myocardium, despite no difference in mitochondrial content. Myocardial tissue from n-3 PUFA patients also displayed greater expression and activity of key antioxidant/anti-inflammatory enzymes. These findings lead to our hypothesis that PPARγ activation is a mechanism by which fish oil n-3 PUFAs enhance mitochondrial fatty acid oxidation and antioxidant capacity in human atrial myocardium, and that this preoperative therapeutic regimen may be optimal for mitigating oxidative/inflammatory stress associated with cardiac surgery.
Figures
References
-
- De Caterina R. n-3 fatty acids in cardiovascular disease. N Engl J Med 364: 2439–2450, 2011 - PubMed
-
- Metcalf RG, James MJ, Gibson RA, Edwards JR, Stubberfield J, Stuklis R, Roberts-Thomson K, Young GD, and Cleland LG. Effects of fish-oil supplementation on myocardial fatty acids in humans. Am J Clin Nutr 85: 1222–1228, 2007 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
