

C57BL/6N albino/agouti mutant mice as embryo donors for efficient germline transmission of C57BL/6 ES cells
- PMID: 24599260
- PMCID: PMC3944090
- DOI: 10.1371/journal.pone.0090570
C57BL/6N albino/agouti mutant mice as embryo donors for efficient germline transmission of C57BL/6 ES cells
Abstract
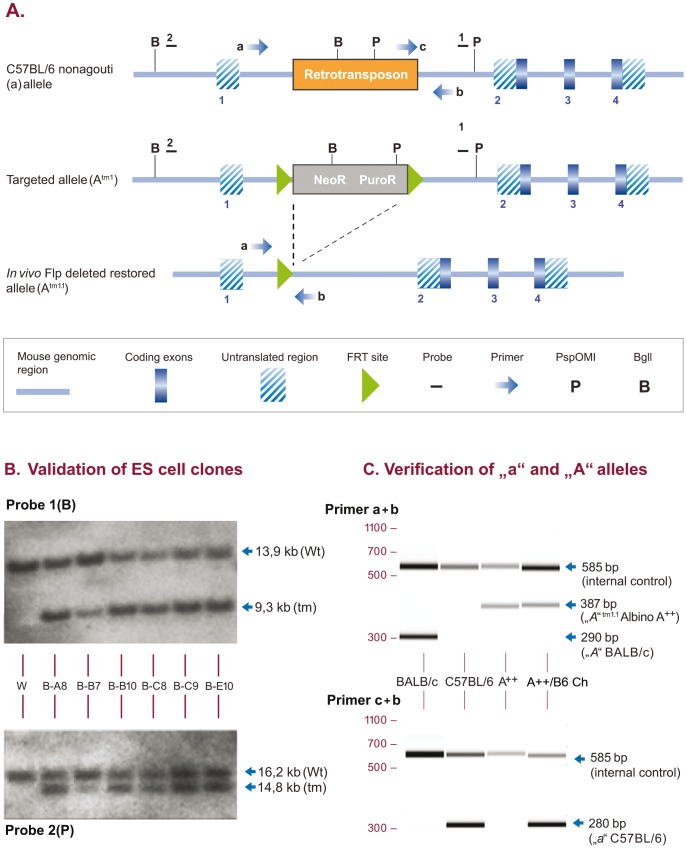
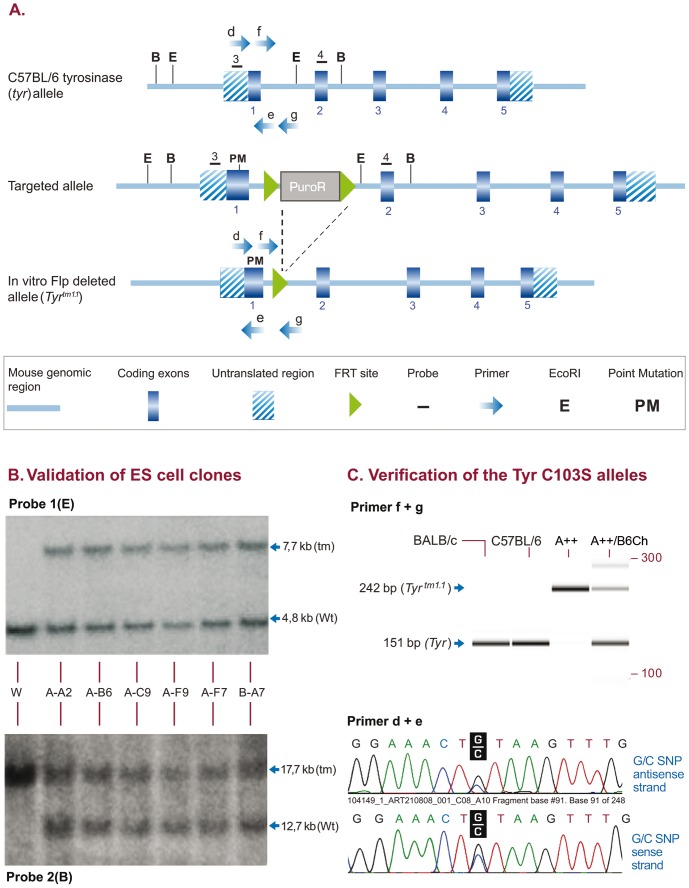
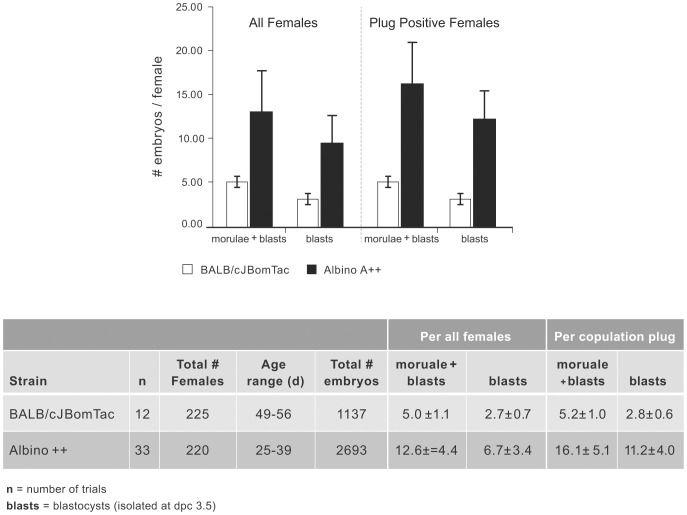
We generated C57BL/6NTac mice carrying a tyrosinase loss-of function mutation and a reversion of the nonagouti locus to agouti. This strain has a high superovulation response, allows visual detection of chimeric coat color contribution of C57BL/6 ES-cells and provides a simplified breeding format that generates black G1 offspring of pure inbred C57BL/6 background in one step, providing the ideal host for genetically manipulated C57BL/6 ES cells.
Conflict of interest statement
Figures

Similar articles
-
Generating Chimeric Mice by Using Embryos from Nonsuperovulated BALB/c Mice Compared with Superovulated BALB/c and Albino C57BL/6 Mice.J Am Assoc Lab Anim Sci. 2016;55(4):400-5. J Am Assoc Lab Anim Sci. 2016. PMID: 27423145 Free PMC article.
-
Agouti C57BL/6N embryonic stem cells for mouse genetic resources.Nat Methods. 2009 Jul;6(7):493-5. doi: 10.1038/nmeth.1342. Epub 2009 Jun 14. Nat Methods. 2009. PMID: 19525957 Free PMC article.
-
A new coat color mouse line for testing germline transmission of embryonic stem cells while retaining an inbred genetic background.J Am Assoc Lab Anim Sci. 2007 May;46(3):37-40. J Am Assoc Lab Anim Sci. 2007. PMID: 17487951
-
Molecular structure of the tyrosinase gene.Pigment Cell Res. 1992;Suppl 2:61-6. doi: 10.1111/j.1600-0749.1990.tb00351.x. Pigment Cell Res. 1992. PMID: 1409440 Review. No abstract available.
-
The molecular basis for dominant yellow agouti coat color mutations.Bioessays. 1994 Oct;16(10):705-7. doi: 10.1002/bies.950161002. Bioessays. 1994. PMID: 7980472 Review.
Cited by
-
Generating Chimeric Mice by Using Embryos from Nonsuperovulated BALB/c Mice Compared with Superovulated BALB/c and Albino C57BL/6 Mice.J Am Assoc Lab Anim Sci. 2016;55(4):400-5. J Am Assoc Lab Anim Sci. 2016. PMID: 27423145 Free PMC article.
-
Comparison of BALB/c and B6-albino mouse strain blastocysts as hosts for the injection of C57BL6/N-derived C2 embryonic stem cells.Transgenic Res. 2016 Aug;25(4):527-31. doi: 10.1007/s11248-016-9937-5. Epub 2016 Feb 6. Transgenic Res. 2016. PMID: 26852382
-
3R measures in facilities for the production of genetically modified rodents.Lab Anim (NY). 2022 Jun;51(6):162-177. doi: 10.1038/s41684-022-00978-1. Epub 2022 May 31. Lab Anim (NY). 2022. PMID: 35641635 Review.
-
Genetic quality: a complex issue for experimental study reproducibility.Transgenic Res. 2022 Oct;31(4-5):413-430. doi: 10.1007/s11248-022-00314-w. Epub 2022 Jun 25. Transgenic Res. 2022. PMID: 35751794 Free PMC article. Review.
-
Efficiency comparison of B6(Cg)-Tyrc-2j /J and C57BL/6NTac embryos as hosts for the generation of knockout mice.Transgenic Res. 2021 Jun;30(3):275-281. doi: 10.1007/s11248-021-00248-9. Epub 2021 Apr 12. Transgenic Res. 2021. PMID: 33844149 Free PMC article.
References
-
- Collins FS, Rossant J, Wurst W (2007) A mouse for all reasons. Cell 128: 9–13. - PubMed
-
- Ledermann B, Burki K (1991) Establishment of a germ-line competent C57BL/6 embryonic stem cell line. Experimental Cell Research 197: 254–258. - PubMed
-
- Schuster-Gossler K, Lee AW, Lerner CP, Parker HJ, Dyer VW, et al.. (2001) Use of coisogenic host blastocysts for efficient establishment of germline chimeras with C57BL/6J ES cell lines. Biotechniques 31: : 1022–1024, 1026. - PubMed
-
- Cheng J, Dutra A, Takesono A, Garrett-Beal L, Schwartzberg PL (2004) Improved generation of C57BL/6J mouse embryonic stem cells in a defined serum-free media. Genesis 39: 100–104. - PubMed
-
- Kontgen F, Suss G, Stewart C, Steinmetz M, Bluethmann H (1993) Targeted disruption of the MHC class II Aa gene in C57BL/6 mice. International Immunology 5: 957–964. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
