

Acute suppressive and long-term phase modulation actions of orexin on the mammalian circadian clock
- PMID: 24599460
- PMCID: PMC3942578
- DOI: 10.1523/JNEUROSCI.3388-13.2014
Acute suppressive and long-term phase modulation actions of orexin on the mammalian circadian clock
Abstract
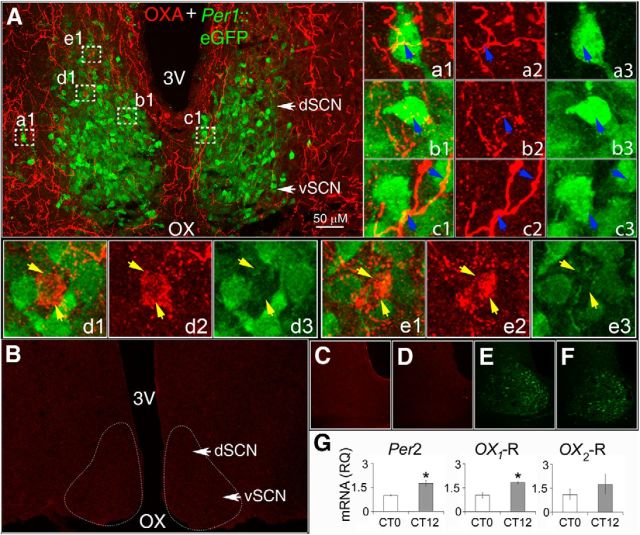
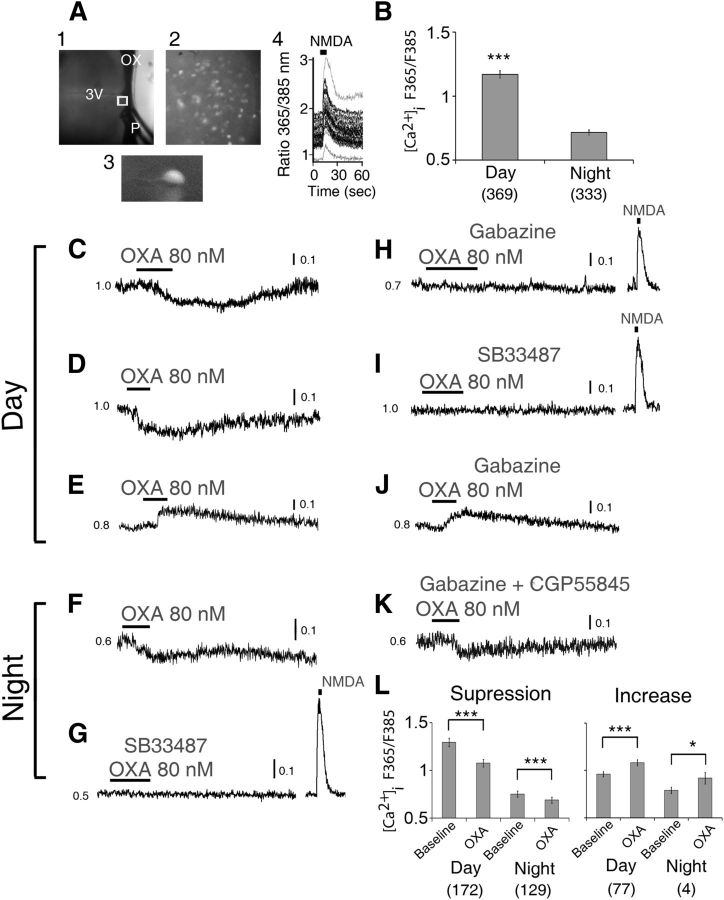
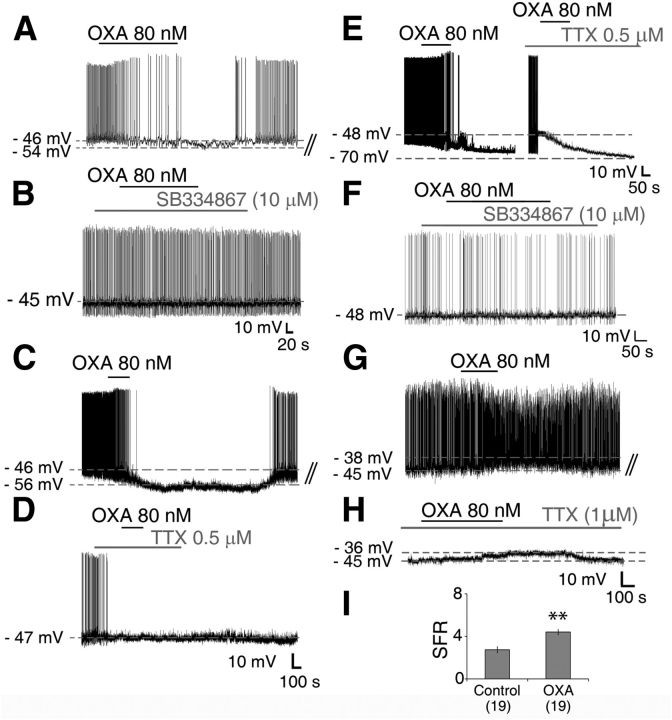
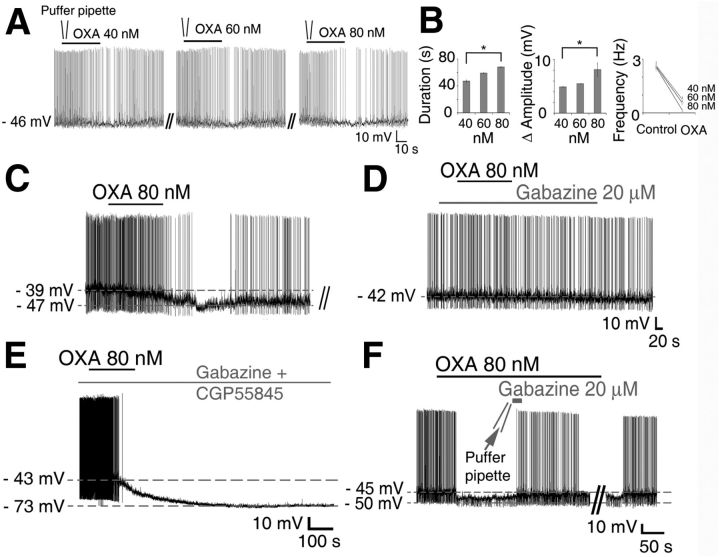
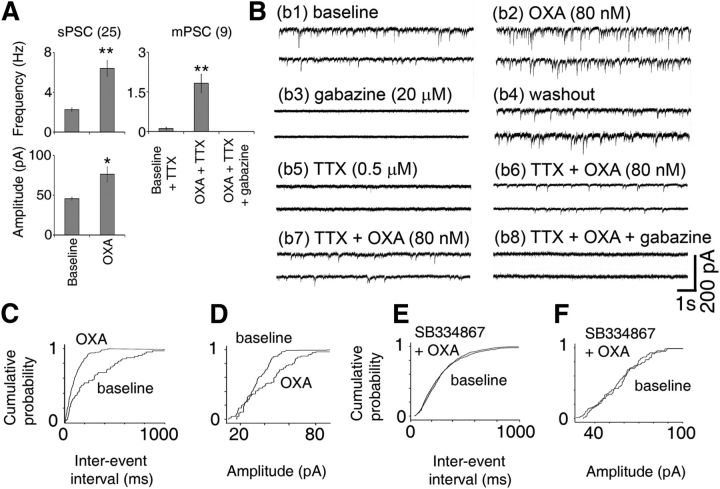
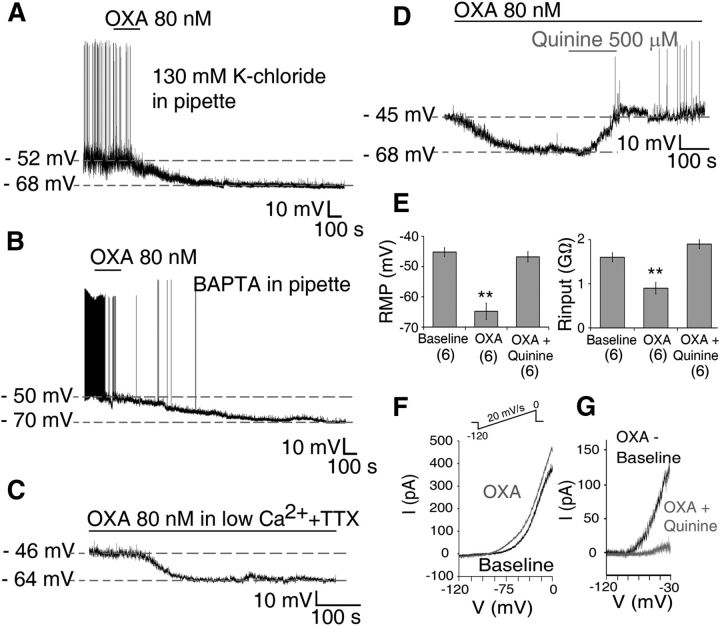
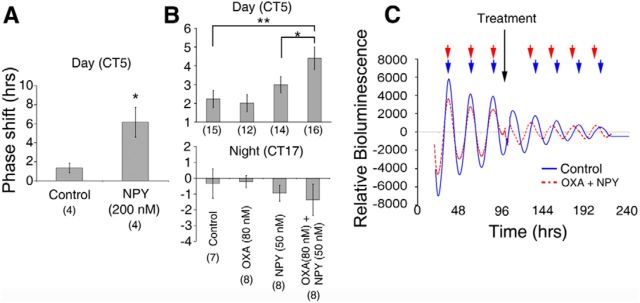
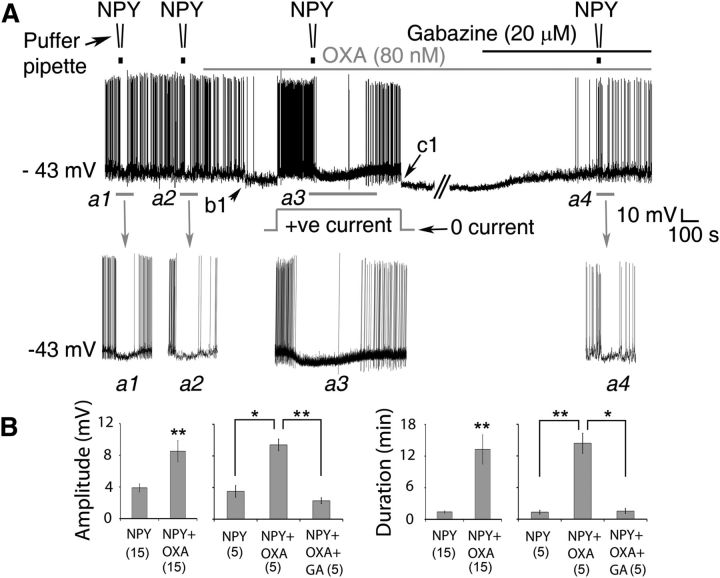
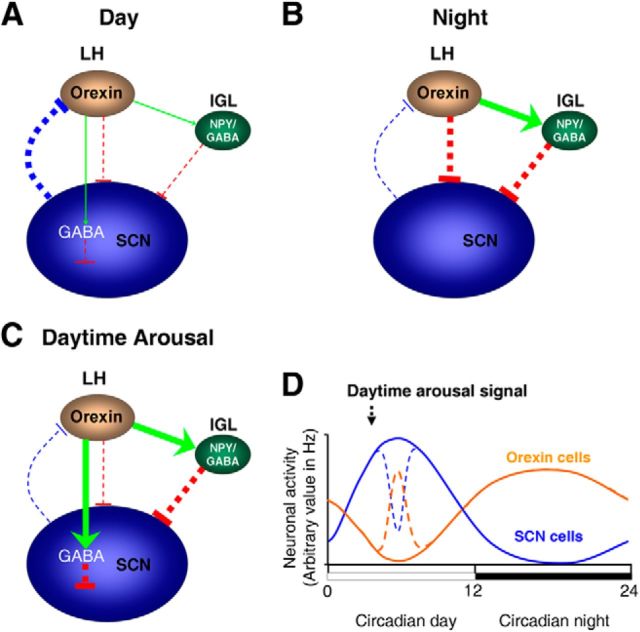
Circadian and homeostatic neural circuits organize the temporal architecture of physiology and behavior, but knowledge of their interactions is imperfect. For example, neurons containing the neuropeptide orexin homeostatically control arousal and appetitive states, while neurons in the suprachiasmatic nuclei (SCN) function as the brain's master circadian clock. The SCN regulates orexin neurons so that they are much more active during the circadian night than the circadian day, but it is unclear whether the orexin neurons reciprocally regulate the SCN clock. Here we show both orexinergic innervation and expression of genes encoding orexin receptors (OX1 and OX2) in the mouse SCN, with OX1 being upregulated at dusk. Remarkably, we find through in vitro physiological recordings that orexin predominantly suppresses mouse SCN Period1 (Per1)-EGFP-expressing clock cells. The mechanisms underpinning these suppressions vary across the circadian cycle, from presynaptic modulation of inhibitory GABAergic signaling during the day to directly activating leak K(+) currents at night. Orexin also augments the SCN clock-resetting effects of neuropeptide Y (NPY), another neurochemical correlate of arousal, and potentiates NPY's inhibition of SCN Per1-EGFP cells. These results build on emerging literature that challenge the widely held view that orexin signaling is exclusively excitatory and suggest new mechanisms for avoiding conflicts between circadian clock signals and homeostatic cues in the brain.
Keywords: GABA; arousal; circadian; electrophysiology; orexin A; suprachiasmatic.
Figures
References
-
- Briston SJ, Caldwell JL, Horn MA, Clarke JD, Richards MA, Greensmith DJ, Graham HK, Hall MC, Eisner DA, Dibb KM, Trafford AW. Impaired beta-adrenergic responsiveness accentuates dysfunctional excitation-contraction coupling in an ovine model of tachypacing-induced heart failure. J Physiol. 2011;589:1367–1382. doi: 10.1113/jphysiol.2010.203984. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous