

Heteromeric Kv7.2/7.3 channels differentially regulate action potential initiation and conduction in neocortical myelinated axons
- PMID: 24599470
- PMCID: PMC3942587
- DOI: 10.1523/JNEUROSCI.4206-13.2014
Heteromeric Kv7.2/7.3 channels differentially regulate action potential initiation and conduction in neocortical myelinated axons
Abstract
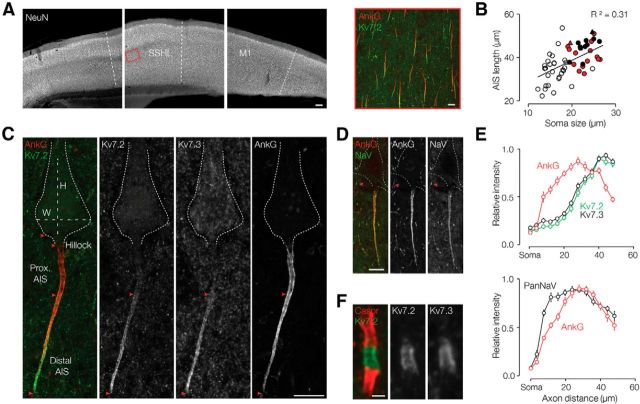
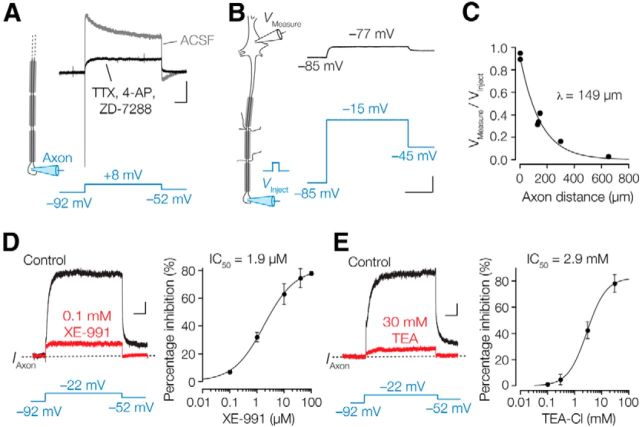
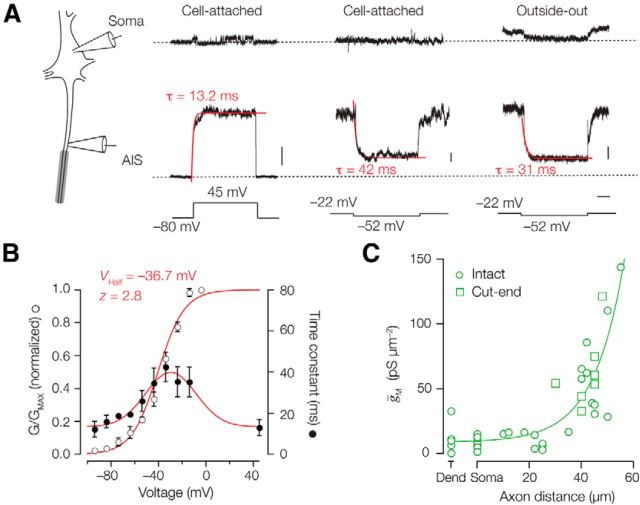
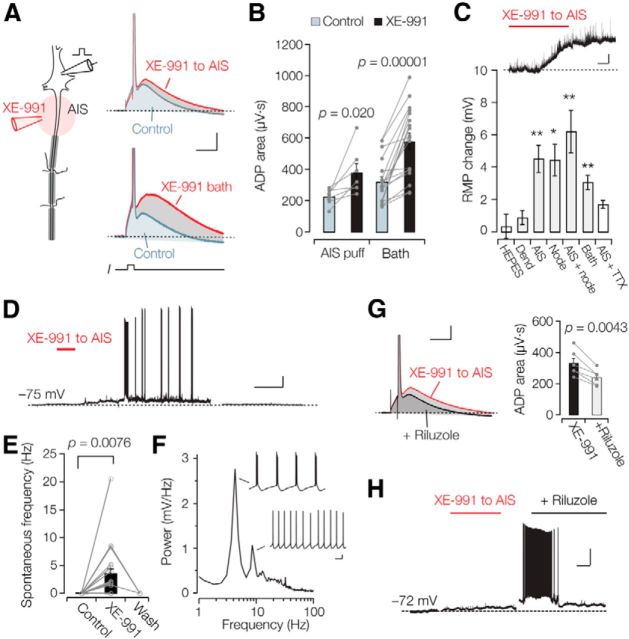
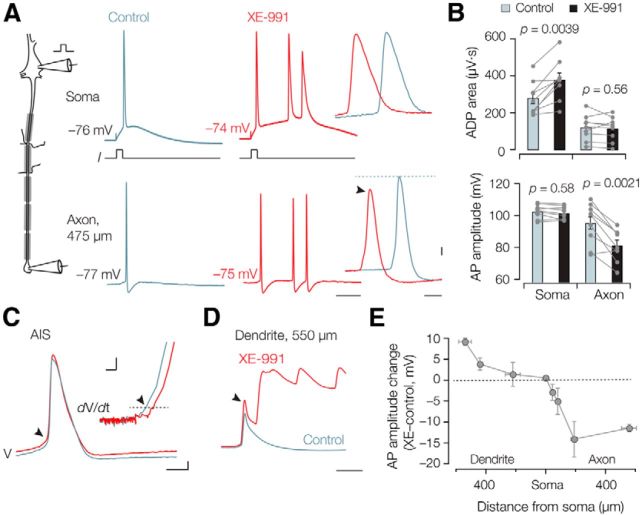
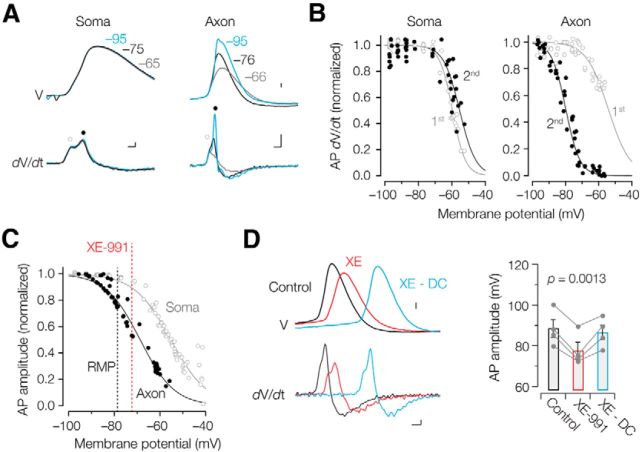
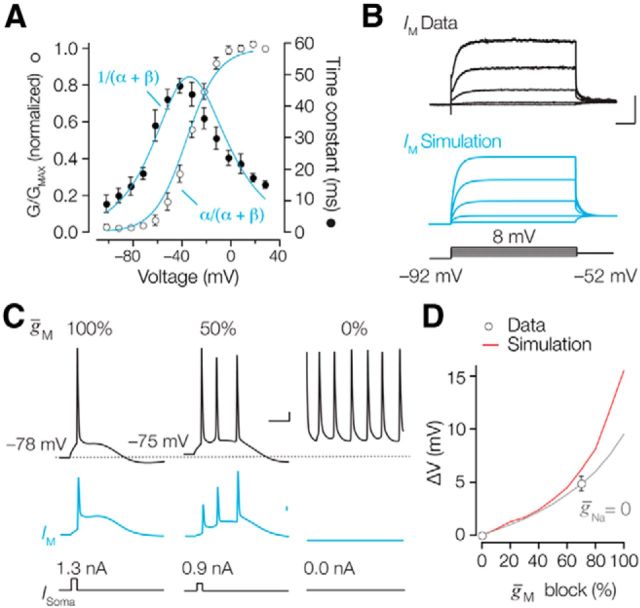
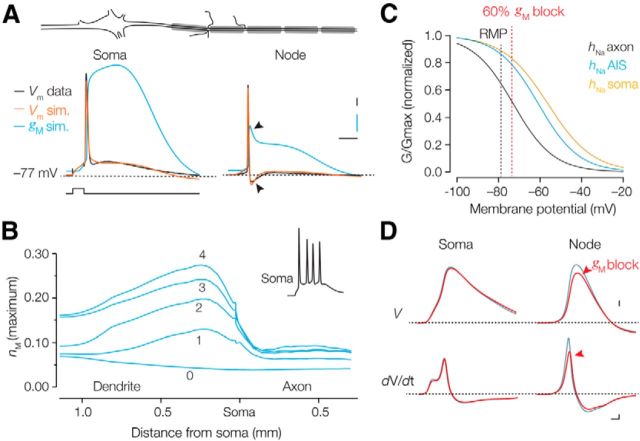
Rapid energy-efficient signaling along vertebrate axons is achieved through intricate subcellular arrangements of voltage-gated ion channels and myelination. One recently appreciated example is the tight colocalization of K(v)7 potassium channels and voltage-gated sodium (Na(v)) channels in the axonal initial segment and nodes of Ranvier. The local biophysical properties of these K(v)7 channels and the functional impact of colocalization with Na(v) channels remain poorly understood. Here, we quantitatively examined K(v)7 channels in myelinated axons of rat neocortical pyramidal neurons using high-resolution confocal imaging and patch-clamp recording. K(v)7.2 and 7.3 immunoreactivity steeply increased within the distal two-thirds of the axon initial segment and was mirrored by the conductance density estimates, which increased from ~12 (proximal) to 150 pS μm(-2) (distal). The axonal initial segment and nodal M-currents were similar in voltage dependence and kinetics, carried by K(v)7.2/7.3 heterotetramers, 4% activated at the resting membrane potential and rapidly activated with single-exponential time constants (~15 ms at 28 mV). Experiments and computational modeling showed that while somatodendritic K(v)7 channels are strongly activated by the backpropagating action potential to attenuate the afterdepolarization and repetitive firing, axonal K(v)7 channels are minimally recruited by the forward-propagating action potential. Instead, in nodal domains K(v)7.2/7.3 channels were found to increase Na(v) channel availability and action potential amplitude by stabilizing the resting membrane potential. Thus, K(v)7 clustering near axonal Na(v) channels serves specific and context-dependent roles, both restraining initiation and enhancing conduction of the action potential.
Keywords: Kv7; axon; excitability.
Figures


Comment in
-
Axonal Kv7.2/7.3 channels: caught in the act.Channels (Austin). 2014;8(4):288-9. doi: 10.4161/chan.29965. Channels (Austin). 2014. PMID: 25171180 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources