

Higher-order structure of the Rous sarcoma virus SP assembly domain
- PMID: 24599998
- PMCID: PMC4019137
- DOI: 10.1128/JVI.02659-13
Higher-order structure of the Rous sarcoma virus SP assembly domain
Abstract
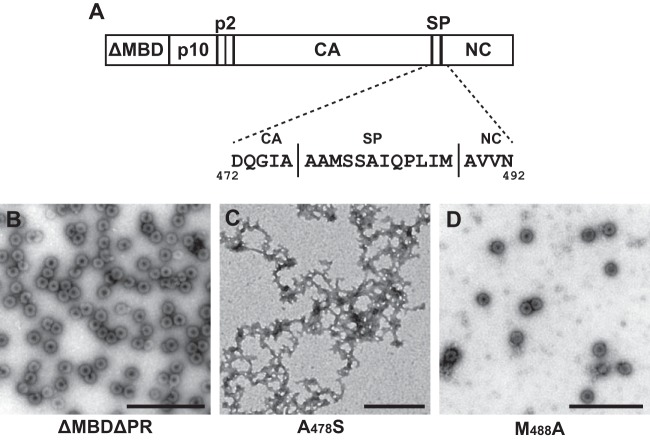
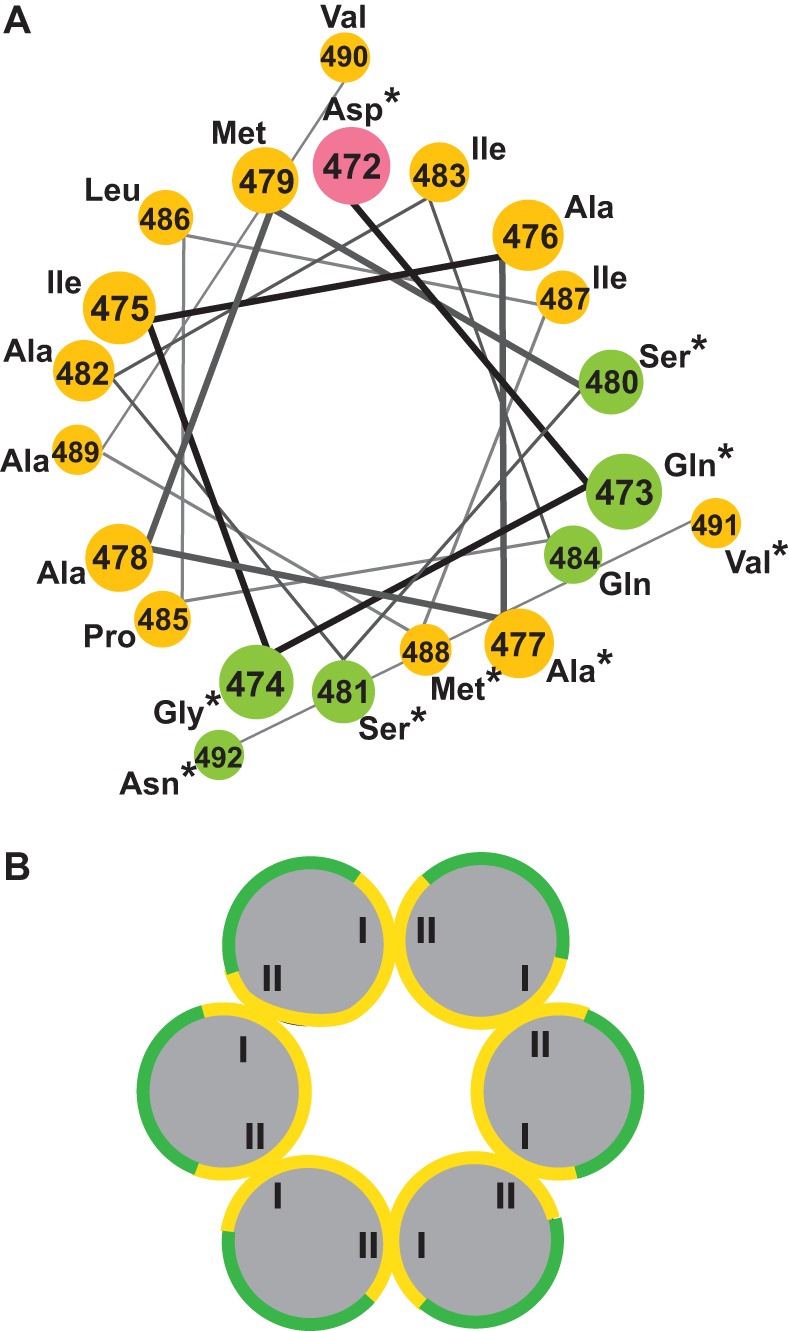
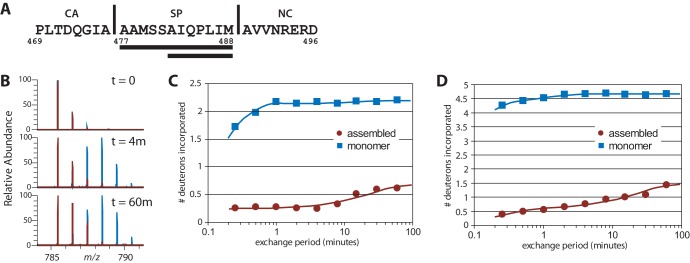
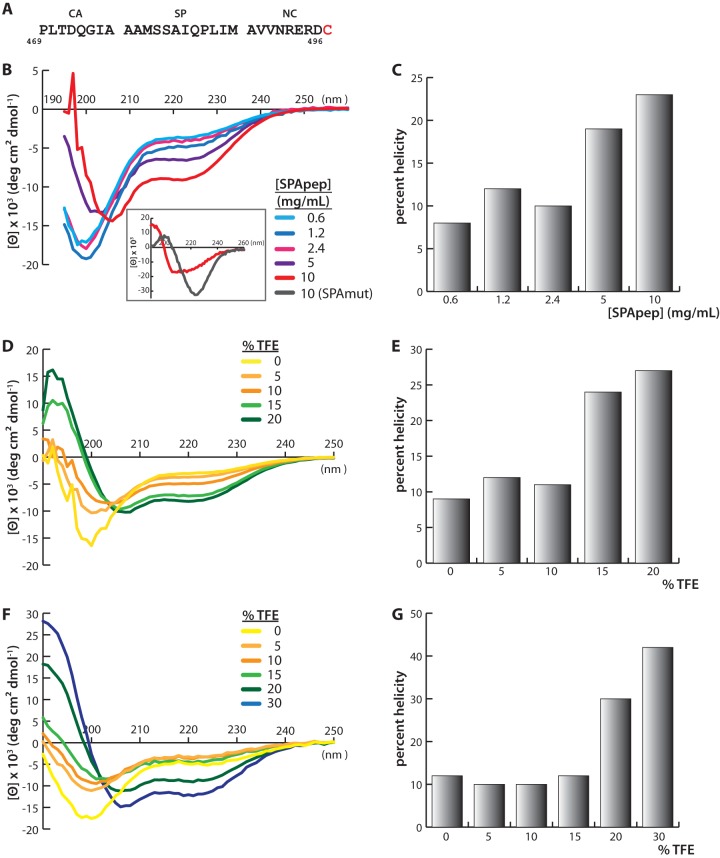
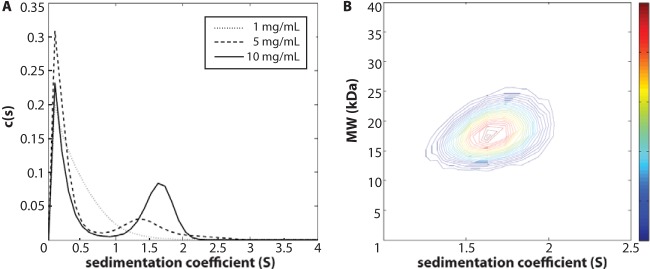
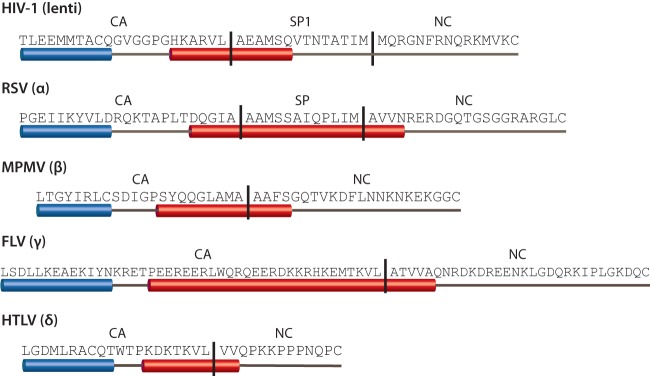
Purified retroviral Gag proteins can assemble in vitro to form immature virus-like particles (VLPs). By cryoelectron tomography, Rous sarcoma virus VLPs show an organized hexameric lattice consisting chiefly of the capsid (CA) domain, with periodic stalk-like densities below the lattice. We hypothesize that the structure represented by these densities is formed by amino acid residues immediately downstream of the folded CA, namely, the short spacer peptide SP, along with a dozen flanking residues. These 24 residues comprise the SP assembly (SPA) domain, and we propose that neighboring SPA units in a Gag hexamer coalesce to form a six-helix bundle. Using in vitro assembly, alanine scanning mutagenesis, and biophysical analyses, we have further characterized the structure and function of SPA. Most of the amino acid residues in SPA could not be mutated individually without abrogating assembly, with the exception of a few residues near the N and C termini, as well as three hydrophilic residues within SPA. We interpret these results to mean that the amino acids that do not tolerate mutations contribute to higher-order structures in VLPs. Hydrogen-deuterium exchange analyses of unassembled Gag compared that of assembled VLPs showed strong protection at the SPA region, consistent with a higher-order structure. Circular dichroism revealed that a 29mer SPA peptide shifts from a random coil to a helix in a concentration-dependent manner. Analytical ultracentrifugation showed concentration-dependent self-association of the peptide into a hexamer. Taken together, these results provide strong evidence for the formation of a critical six-helix bundle in Gag assembly.
Importance: The structure of a retrovirus like HIV is created by several thousand molecules of the viral Gag protein, which assemble to form the known hexagonal protein lattice in the virus particle. How the Gag proteins pack together in the lattice is incompletely understood. A short segment of Gag known to be critical for proper assembly has been hypothesized to form a six-helix bundle, which may be the nucleating event that leads to lattice formation. The experiments reported here, using the avian Rous sarcoma virus as a model system, further define the nature of this segment of Gag, show that it is in a higher-order structure in the virus particle, and provide the first direct evidence that it forms a six-helix bundle in retrovirus assembly. Such knowledge may provide underpinnings for the development of antiretroviral drugs that interfere with virus assembly.
Figures

Similar articles
-
Membrane Binding of the Rous Sarcoma Virus Gag Protein Is Cooperative and Dependent on the Spacer Peptide Assembly Domain.J Virol. 2015 Dec 16;90(5):2473-85. doi: 10.1128/JVI.02733-15. J Virol. 2015. PMID: 26676779 Free PMC article.
-
Mutations in the spacer peptide and adjoining sequences in Rous sarcoma virus Gag lead to tubular budding.J Virol. 2008 Jul;82(14):6788-97. doi: 10.1128/JVI.00213-08. Epub 2008 Apr 30. J Virol. 2008. PMID: 18448521 Free PMC article.
-
The Structure of Immature Virus-Like Rous Sarcoma Virus Gag Particles Reveals a Structural Role for the p10 Domain in Assembly.J Virol. 2015 Oct;89(20):10294-302. doi: 10.1128/JVI.01502-15. Epub 2015 Jul 29. J Virol. 2015. PMID: 26223638 Free PMC article.
-
A Structural Perspective of the Role of IP6 in Immature and Mature Retroviral Assembly.Viruses. 2021 Sep 17;13(9):1853. doi: 10.3390/v13091853. Viruses. 2021. PMID: 34578434 Free PMC article. Review.
-
Membrane interaction of retroviral Gag proteins.Front Microbiol. 2014 Apr 29;5:187. doi: 10.3389/fmicb.2014.00187. eCollection 2014. Front Microbiol. 2014. PMID: 24808894 Free PMC article. Review.
Cited by
-
Computational Methodologies for Real-Space Structural Refinement of Large Macromolecular Complexes.Annu Rev Biophys. 2016 Jul 5;45:253-78. doi: 10.1146/annurev-biophys-062215-011113. Epub 2016 May 2. Annu Rev Biophys. 2016. PMID: 27145875 Free PMC article. Review.
-
Effect of Small Polyanions on In Vitro Assembly of Selected Members of Alpha-, Beta- and Gammaretroviruses.Viruses. 2021 Jan 18;13(1):129. doi: 10.3390/v13010129. Viruses. 2021. PMID: 33477490 Free PMC article.
-
Potential role for CA-SP in nucleating retroviral capsid maturation.J Virol. 2014 Jul;88(13):7170-7. doi: 10.1128/JVI.00309-14. Epub 2014 Apr 9. J Virol. 2014. PMID: 24719425 Free PMC article.
-
Atomic Modeling of an Immature Retroviral Lattice Using Molecular Dynamics and Mutagenesis.Structure. 2015 Aug 4;23(8):1414-1425. doi: 10.1016/j.str.2015.05.017. Epub 2015 Jun 25. Structure. 2015. PMID: 26118533 Free PMC article.
-
Role of Mason-Pfizer monkey virus CA-NC spacer peptide-like domain in assembly of immature particles.J Virol. 2014 Dec;88(24):14148-60. doi: 10.1128/JVI.02286-14. Epub 2014 Oct 1. J Virol. 2014. PMID: 25275119 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
