

Ribosome biogenesis: emerging evidence for a central role in the regulation of skeletal muscle mass
- PMID: 24604615
- PMCID: PMC4868551
- DOI: 10.1002/jcp.24604
Ribosome biogenesis: emerging evidence for a central role in the regulation of skeletal muscle mass
Abstract
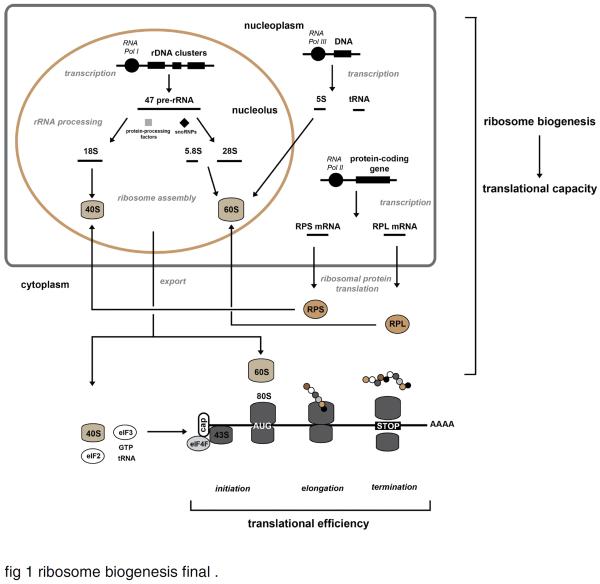
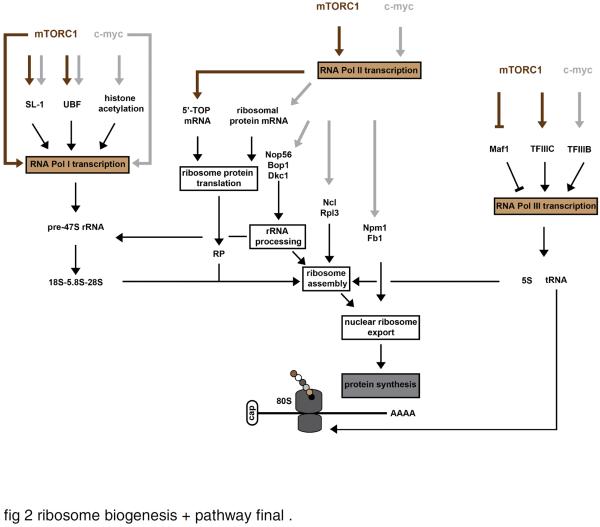
The ribosome is a supramolecular ribonucleoprotein complex that functions at the heart of the translation machinery to convert mRNA into protein. Ribosome biogenesis is the primary determinant of translational capacity of the cell and accordingly has an essential role in the control of cell growth in eukaryotes. Cumulative evidence supports the hypothesis that ribosome biogenesis has an important role in the regulation of skeletal muscle mass. The purpose of this review is to, first, summarize the main mechanisms known to regulate ribosome biogenesis and, second, put forth the hypothesis that ribosome biogenesis is a central mechanism used by skeletal muscle to regulate protein synthesis and control skeletal muscle mass in response to anabolic and catabolic stimuli. The mTORC1 and Wnt/β-catenin/c-myc signaling pathways are discussed as the major pathways that work in concert with each of the three RNA polymerases (RNA Pol I, II, and III) in regulating ribosome biogenesis. Consistent with our hypothesis, activation of these two pathways has been shown to be associated with ribosome biogenesis during skeletal muscle hypertrophy. Although further study is required, the finding that ribosome biogenesis is altered under catabolic states, in particular during disuse atrophy, suggests that its activation represents a novel therapeutic target to reduce or prevent muscle atrophy. Lastly, the emerging field of ribosome specialization is discussed and its potential role in the regulation of gene expression during periods of skeletal muscle plasticity.
© 2014 Wiley Periodicals, Inc.
Figures
References
-
- Adams GR, Caiozzo VJ, Haddad F, Baldwin KM. Cellular and molecular responses to increased skeletal muscle loading after irradiation. Am J Physiol Cell Physiol. 2002;283(4):C1182–1195. - PubMed
-
- Alawi F, Lee MN. DKC1 is a direct and conserved transcriptional target of c-MYC. Biochem Biophys Res Commun. 2007;362(4):893–898. - PubMed
-
- Arabi A, Wu S, Ridderstrale K, Bierhoff H, Shiue C, Fatyol K, Fahlen S, Hydbring P, Soderberg O, Grummt I, Larsson LG, Wright AP. c-Myc associates with ribosomal DNA and activates RNA polymerase I transcription. Nat Cell Biol. 2005;7(3):303–310. - PubMed
-
- Armstrong DD, Esser KA. Wnt/beta-catenin signaling activates growth-control genes during overload-induced skeletal muscle hypertrophy. Am J Physiol Cell Physiol. 2005;289(4):C853–859. - PubMed
-
- Armstrong DD, Wong VL, Esser KA. Expression of beta-catenin is necessary for physiological growth of adult skeletal muscle. Am J Physiol Cell Physiol. 2006;291(1):C185–188. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
