

Peripheral coding of taste
- PMID: 24607224
- PMCID: PMC3994536
- DOI: 10.1016/j.neuron.2014.02.022
Peripheral coding of taste
Abstract
Five canonical tastes, bitter, sweet, umami (amino acid), salty, and sour (acid), are detected by animals as diverse as fruit flies and humans, consistent with a near-universal drive to consume fundamental nutrients and to avoid toxins or other harmful compounds. Surprisingly, despite this strong conservation of basic taste qualities between vertebrates and invertebrates, the receptors and signaling mechanisms that mediate taste in each are highly divergent. The identification over the last two decades of receptors and other molecules that mediate taste has led to stunning advances in our understanding of the basic mechanisms of transduction and coding of information by the gustatory systems of vertebrates and invertebrates. In this Review, we discuss recent advances in taste research, mainly from the fly and mammalian systems, and we highlight principles that are common across species, despite stark differences in receptor types.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
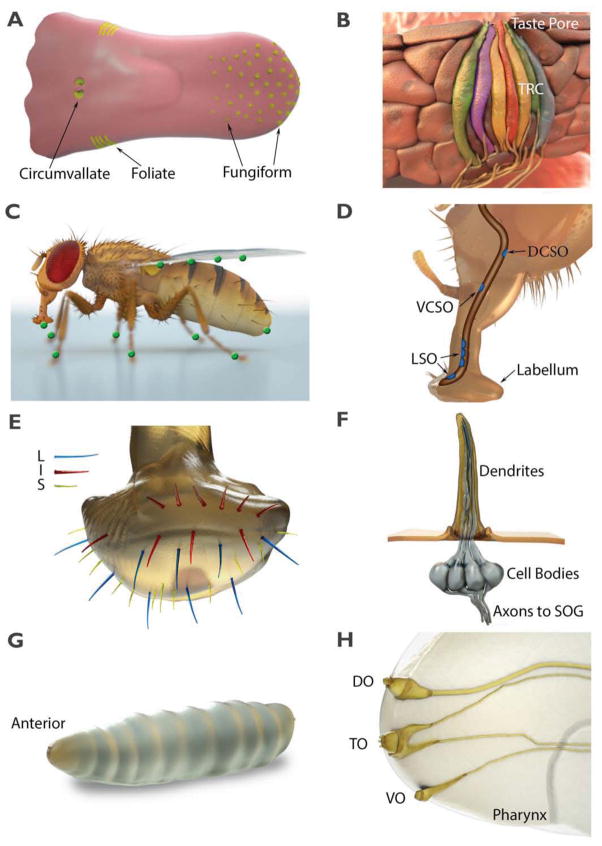
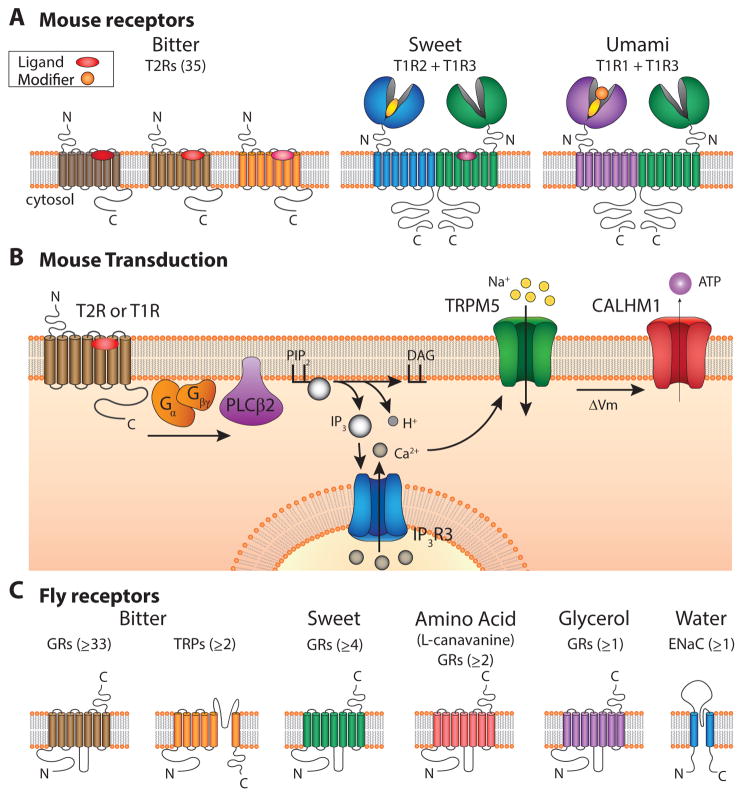
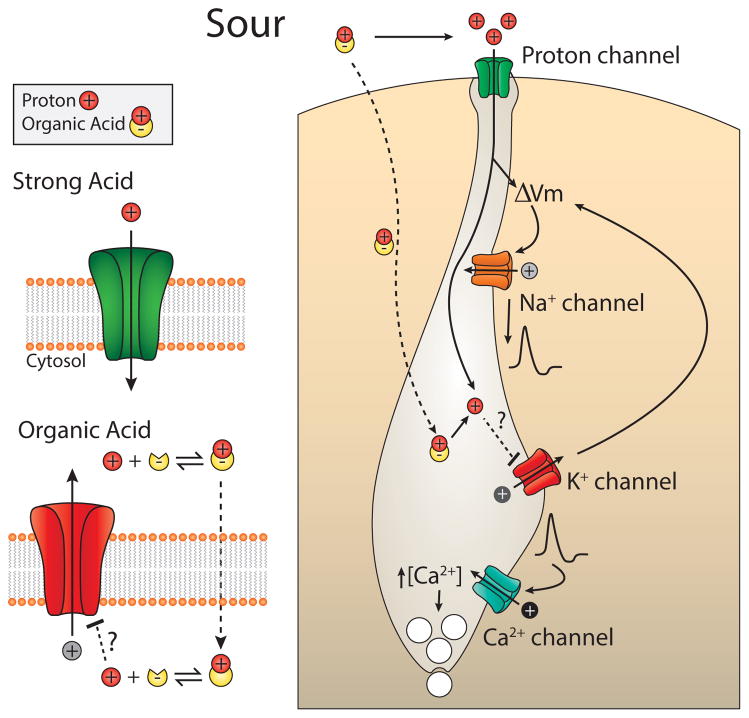
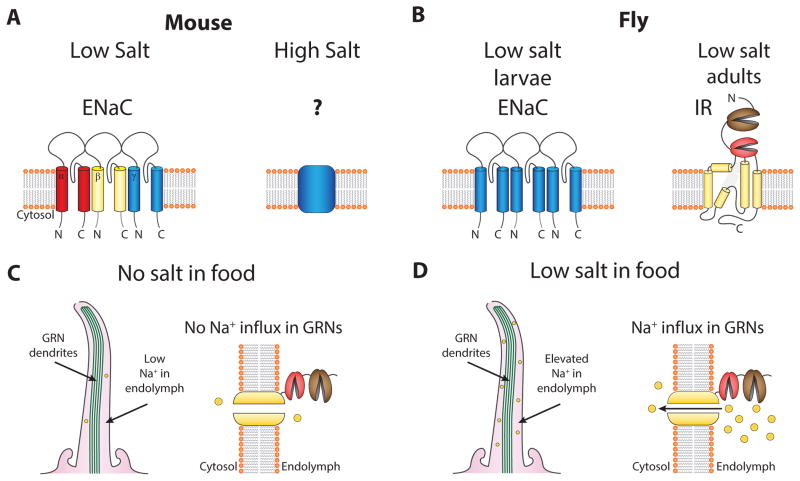
References
-
- Al-Anzi B, Tracey WD, Jr, Benzer S. Response of Drosophila to wasabi is mediated by painless, the fly homolog of mammalian TRPA1/ANKTM1. Curr Biol. 2006;16:1034–1040. - PubMed
-
- Behrens M, Meyerhof W. Mammalian bitter taste perception. Results Probl Cell Differ. 2009;47:203–220. - PubMed
-
- Behrens M, Meyerhof W. Gustatory and extragustatory functions of mammalian taste receptors. Physiol Behav. 2011;105:4–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01EY010852/EY/NEI NIH HHS/United States
- R01 DC007864/DC/NIDCD NIH HHS/United States
- R01DC004564/DC/NIDCD NIH HHS/United States
- R01 DC004564/DC/NIDCD NIH HHS/United States
- R01 EY010852/EY/NEI NIH HHS/United States
- R01DC007864/DC/NIDCD NIH HHS/United States
- R01 EY008117/EY/NEI NIH HHS/United States
- R01 GM085335/GM/NIGMS NIH HHS/United States
- R21DC012747/DC/NIDCD NIH HHS/United States
- R21 DC012747/DC/NIDCD NIH HHS/United States
- R01 DC013741/DC/NIDCD NIH HHS/United States
- R01GM085335/GM/NIGMS NIH HHS/United States
- R01EY008117/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
