

The adaptive trade-off between detection and discrimination in cortical representations and behavior
- PMID: 24607233
- PMCID: PMC4026261
- DOI: 10.1016/j.neuron.2014.01.025
The adaptive trade-off between detection and discrimination in cortical representations and behavior
Abstract
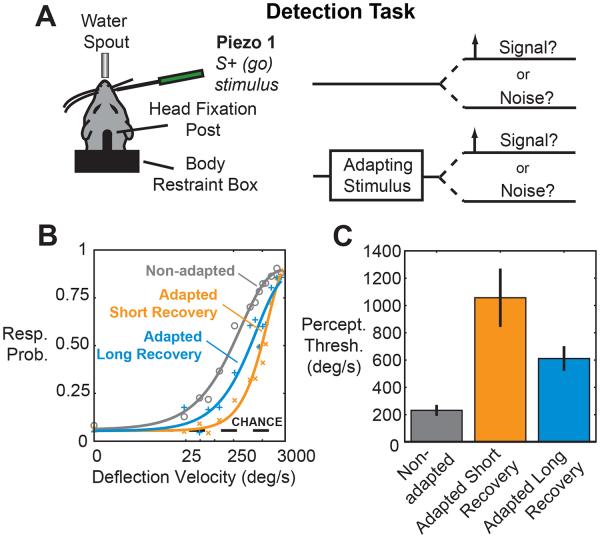
It has long been posited that detectability of sensory inputs can be sacrificed in favor of improved discriminability and that sensory adaptation may mediate this trade-off. The extent to which this trade-off exists behaviorally and the complete picture of the underlying neural representations that likely subserve the phenomenon remain unclear. In the rodent vibrissa system, an ideal observer analysis of cortical activity measured using voltage-sensitive dye imaging in anesthetized animals was combined with behavioral detection and discrimination tasks, thalamic recordings from awake animals, and computational modeling to show that spatial discrimination performance was improved following adaptation, but at the expense of the ability to detect weak stimuli. Together, these results provide direct behavioral evidence for the trade-off between detectability and discriminability, that this trade-off can be modulated through bottom-up sensory adaptation, and that these effects correspond to important changes in thalamocortical coding properties.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures






References
-
- Ahissar E, Sosnik R, Haidarliu S. Transformation from temporal to rate coding in a somatosensory thalamocortical pathway. Nature. 2000;406:302–306. - PubMed
-
- Alonso JM, Usrey WM, Reid RC. Precisely correlated firing in cells of the lateral geniculate nucleus. Nature. 1996;383:815–819. - PubMed
-
- Barlow H. Possible principles underlying the transformation of sensory messages. In Sensory Communication. 1961:271–274.
-
- von Békésy G. Neural volleys and the similarity between some sensations produced by tones and by skin vibrations. The Journal of the Acoustical Society of America. 1957;29:1059–1069.
-
- Brecht M, Preilowski B, Merzenich MM. Functional architecture of the mystacial vibrissae. Behavioural Brain Research. 1997;84:81–97. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
