

Sex differences in Parkinson's disease
- PMID: 24607323
- PMCID: PMC4096384
- DOI: 10.1016/j.yfrne.2014.02.002
Sex differences in Parkinson's disease
Abstract
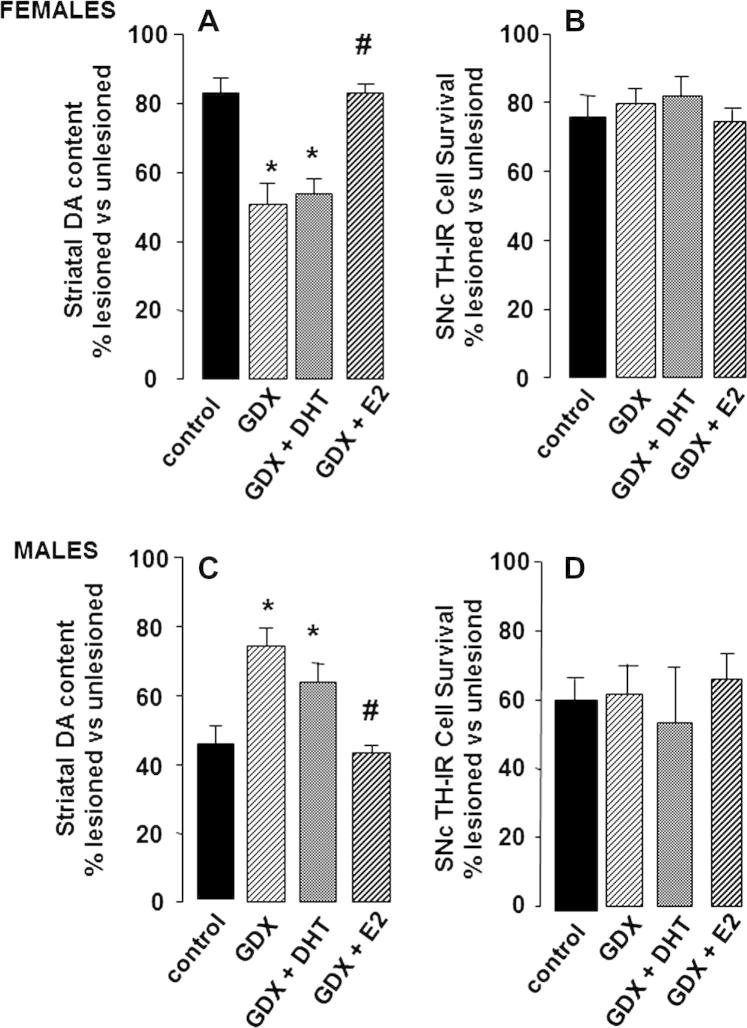
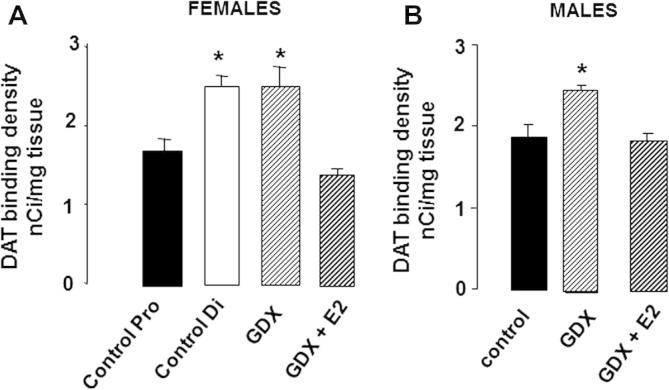
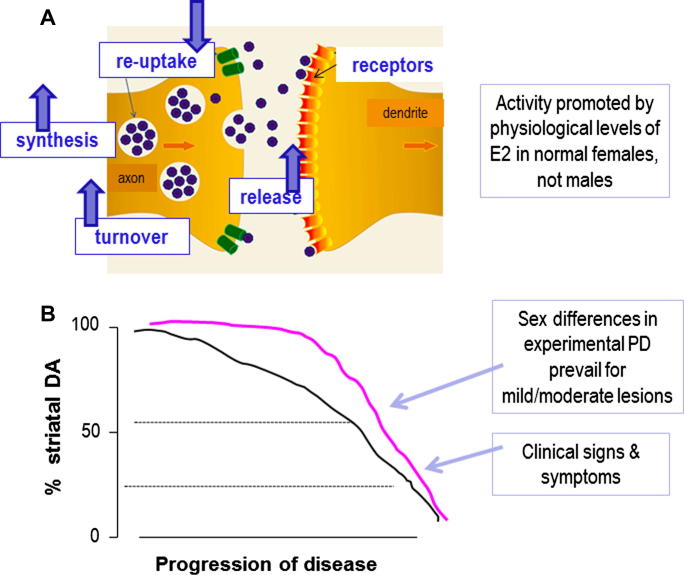
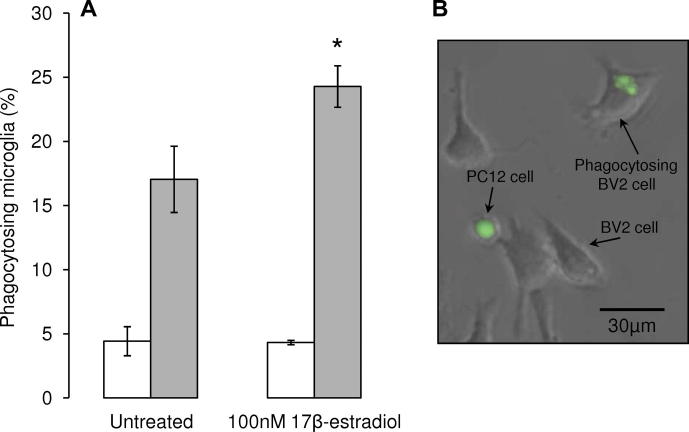
Parkinson's disease (PD) displays a greater prevalence and earlier age at onset in men. This review addresses the concept that sex differences in PD are determined, largely, by biological sex differences in the NSDA system which, in turn, arise from hormonal, genetic and environmental influences. Current therapies for PD rely on dopamine replacement strategies to treat symptoms, and there is an urgent, unmet need for disease modifying agents. As a significant degree of neuroprotection against the early stages of clinical or experimental PD is seen, respectively, in human and rodent females compared with males, a better understanding of brain sex dimorphisms in the intact and injured NSDA system will shed light on mechanisms which have the potential to delay, or even halt, the progression of PD. Available evidence suggests that sex-specific, hormone-based therapeutic agents hold particular promise for developing treatments with optimal efficacy in men and women.
Keywords: Neuroprotection; Nigrostriatal dopaminergic pathways; Parkinson’s disease; Sex differences; Sex hormones.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Abraham I.M., Herbison A.E. Major sex differences in non-genomic estrogen actions on intracellular signaling in mouse brain in vivo. Neuroscience. 2005;131:945–951. - PubMed
-
- Al-Sweidi S., Morissette M., Bourque M., Di Paolo T. Estrogen receptors and gonadal steroids in vulnerability and protection of dopamine neurons in a mouse model of Parkinson’s disease. Neuropharmacology. 2011;61:583–591. - PubMed
-
- Alves G., Muller B., Herlofson K., HogenEsch I., Telstad W., Aarsland D., Tysnes O.B., Larsen J.P. Incidence of Parkinson’s disease in Norway: the Norwegian ParkWest study. J. Neurol. Neurosurg. Psychiatry. 2009;80:851–857. - PubMed
-
- Arevalo M.A., Santos-Galindo M., Acaz-Fonseca E., Azcoitia I., Garcia-Segura L.M. Gonadal hormones and the control of reactive gliosis. Horm. Behav. 2013;63:216–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
