

Modulation of calcium-induced cell death in human neural stem cells by the novel peptidylarginine deiminase-AIF pathway
- PMID: 24607566
- PMCID: PMC3996523
- DOI: 10.1016/j.bbamcr.2014.02.018
Modulation of calcium-induced cell death in human neural stem cells by the novel peptidylarginine deiminase-AIF pathway
Abstract
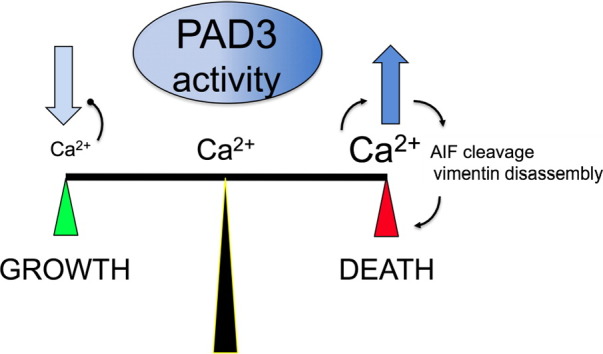
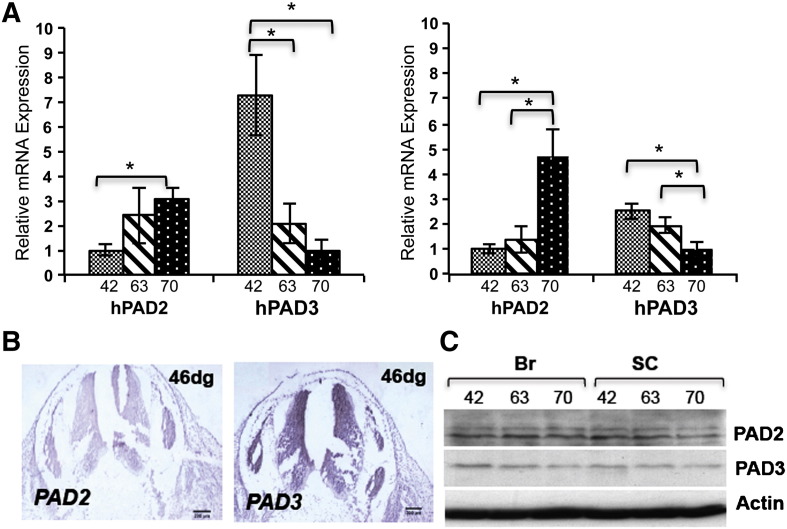
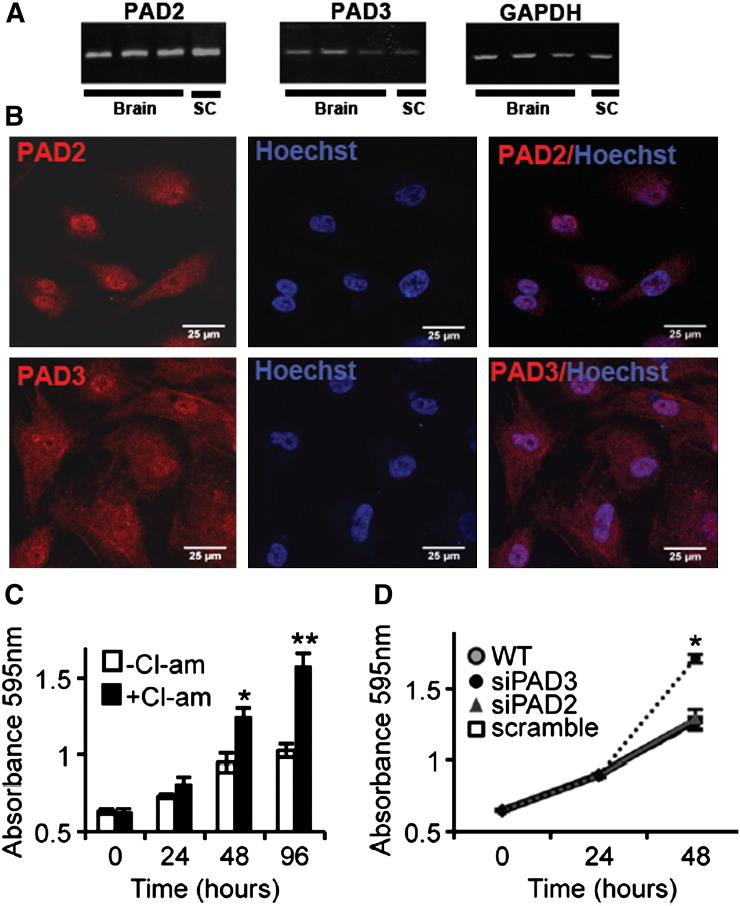
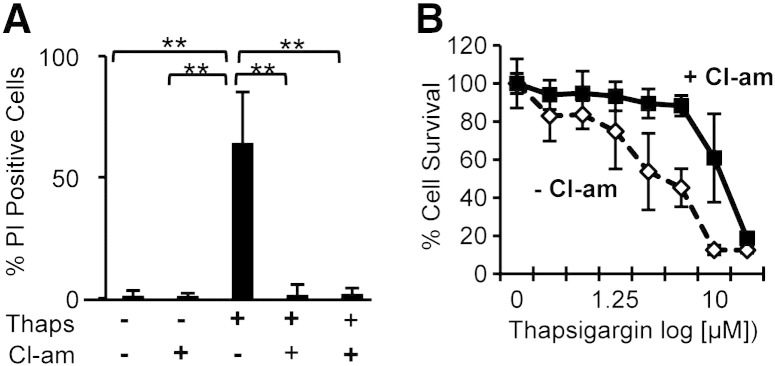
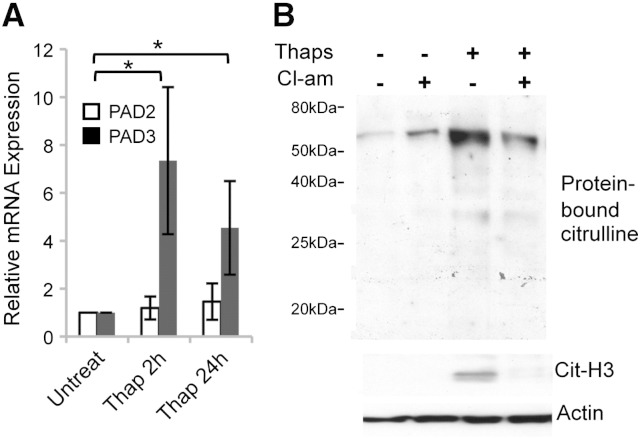
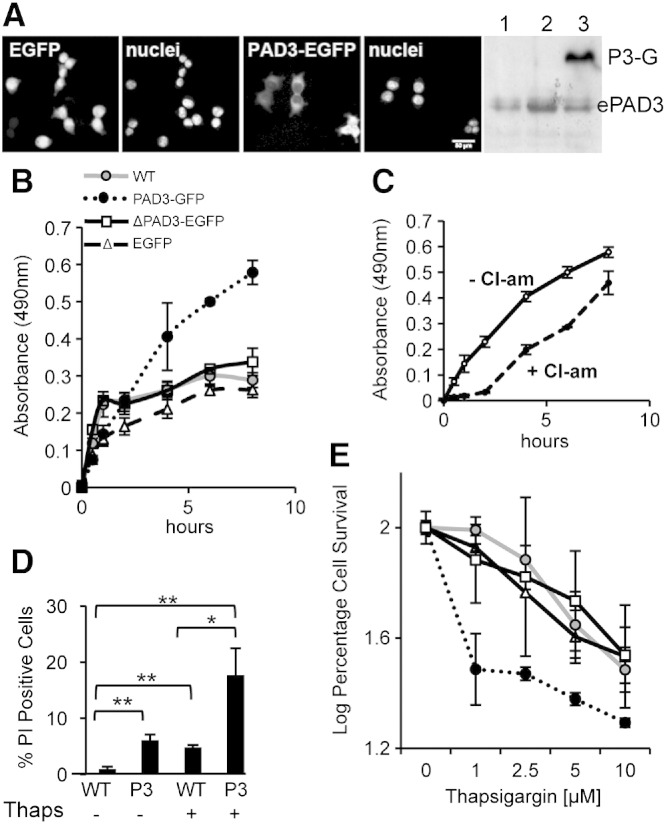
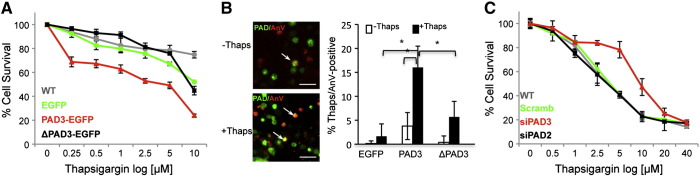
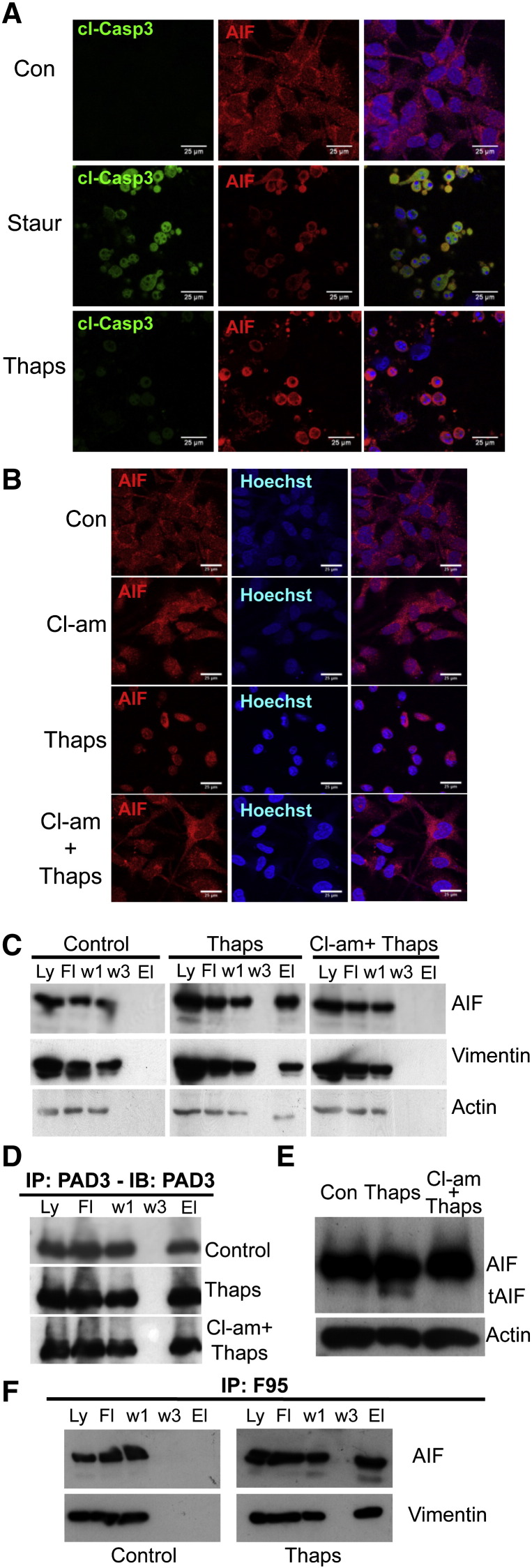
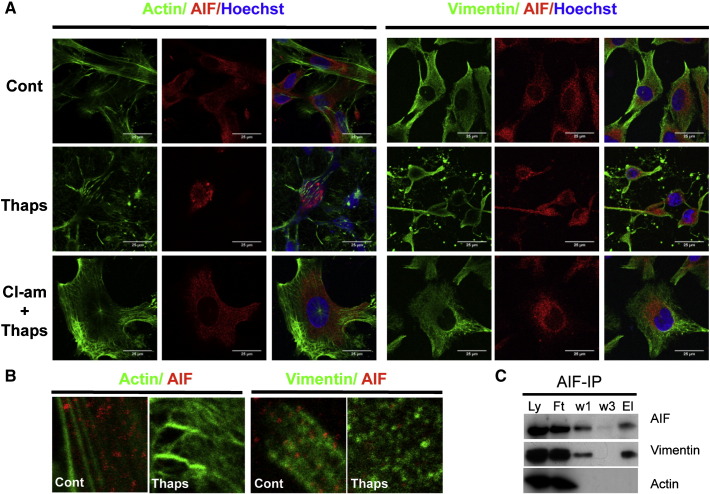
PADs (peptidylarginine deiminases) are calcium-dependent enzymes that change protein-bound arginine to citrulline (citrullination/deimination) affecting protein conformation and function. PAD up-regulation following chick spinal cord injury has been linked to extensive tissue damage and loss of regenerative capability. Having found that human neural stem cells (hNSCs) expressed PAD2 and PAD3, we studied PAD function in these cells and investigated PAD3 as a potential target for neuroprotection by mimicking calcium-induced secondary injury responses. We show that PAD3, rather than PAD2 is a modulator of cell growth/death and that PAD activity is not associated with caspase-3-dependent cell death, but is required for AIF (apoptosis inducing factor)-mediated apoptosis. PAD inhibition prevents association of PAD3 with AIF and AIF cleavage required for its translocation to the nucleus. Finally, PAD inhibition also hinders calcium-induced cytoskeleton disassembly and association of PAD3 with vimentin, that we show to be associated also with AIF; together this suggests that PAD-dependent cytoskeleton disassembly may play a role in AIF translocation to the nucleus. This is the first study highlighting a role of PAD activity in balancing hNSC survival/death, identifying PAD3 as an important upstream regulator of calcium-induced apoptosis, which could be targeted to reduce neural loss, and shedding light on the mechanisms involved.
Keywords: Apoptosis inducing factor (AIF); Cell death; Citrullination–deimination; Human neural stem cell; Peptidylarginine deiminase (PAD, PADI); Vimentin.
Copyright © 2014 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Protein deiminases: new players in the developmentally regulated loss of neural regenerative ability.Dev Biol. 2011 Jul 15;355(2):205-14. doi: 10.1016/j.ydbio.2011.04.015. Epub 2011 Apr 22. Dev Biol. 2011. PMID: 21539830 Free PMC article.
-
Peptidylarginine Deiminase Inhibitor Application, Using Cl-Amidine, PAD2, PAD3 and PAD4 Isozyme-Specific Inhibitors in Pancreatic Cancer Cells, Reveals Roles for PAD2 and PAD3 in Cancer Invasion and Modulation of Extracellular Vesicle Signatures.Int J Mol Sci. 2021 Jan 30;22(3):1396. doi: 10.3390/ijms22031396. Int J Mol Sci. 2021. PMID: 33573274 Free PMC article.
-
Three isozymes of peptidylarginine deiminase in the chicken: molecular cloning, characterization, and tissue distribution.Comp Biochem Physiol B Biochem Mol Biol. 2014 Jan;167:65-73. doi: 10.1016/j.cbpb.2013.10.003. Epub 2013 Oct 23. Comp Biochem Physiol B Biochem Mol Biol. 2014. PMID: 24161753
-
An interplay of structure and intrinsic disorder in the functionality of peptidylarginine deiminases, a family of key autoimmunity-related enzymes.Cell Mol Life Sci. 2019 Dec;76(23):4635-4662. doi: 10.1007/s00018-019-03237-8. Epub 2019 Jul 24. Cell Mol Life Sci. 2019. PMID: 31342121 Free PMC article. Review.
-
Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis.Biochim Biophys Acta. 2013 Oct;1829(10):1126-35. doi: 10.1016/j.bbagrm.2013.07.003. Epub 2013 Jul 13. Biochim Biophys Acta. 2013. PMID: 23860259 Free PMC article. Review.
Cited by
-
Peptidylarginine Deiminases-Roles in Cancer and Neurodegeneration and Possible Avenues for Therapeutic Intervention via Modulation of Exosome and Microvesicle (EMV) Release?Int J Mol Sci. 2017 Jun 5;18(6):1196. doi: 10.3390/ijms18061196. Int J Mol Sci. 2017. PMID: 28587234 Free PMC article. Review.
-
Role of the PADI family in inflammatory autoimmune diseases and cancers: A systematic review.Front Immunol. 2023 Mar 20;14:1115794. doi: 10.3389/fimmu.2023.1115794. eCollection 2023. Front Immunol. 2023. PMID: 37020554 Free PMC article.
-
An Overview of the Intrinsic Role of Citrullination in Autoimmune Disorders.J Immunol Res. 2019 Nov 25;2019:7592851. doi: 10.1155/2019/7592851. eCollection 2019. J Immunol Res. 2019. PMID: 31886309 Free PMC article. Review.
-
K9(C4H4FN2O2)2Nd(PW11O39)2·25H2O induces apoptosis in human lung cancer A549 cells.Oncol Lett. 2017 Mar;13(3):1348-1352. doi: 10.3892/ol.2016.5543. Epub 2016 Dec 28. Oncol Lett. 2017. PMID: 28454260 Free PMC article.
-
Citrullination and PAD Enzyme Biology in Type 1 Diabetes - Regulators of Inflammation, Autoimmunity, and Pathology.Front Immunol. 2021 Jun 1;12:678953. doi: 10.3389/fimmu.2021.678953. eCollection 2021. Front Immunol. 2021. PMID: 34140951 Free PMC article. Review.
References
-
- Jang B., Jin J.K., Jeon Y.C., Cho H.J., Ishigami A., Choi K.C., Carp R.I., Maruyama N., Kim Y.S., Choi E.K. Involvement of peptidylarginine deiminase-mediated post-translational citrullination in pathogenesis of sporadic Creutzfeldt–Jakob disease. Acta Neuropathol. 2010;119:199–210. - PubMed
-
- Jang B., Jeon Y.C., Choi J.K., Park M., Kim J.I., Ishigami A., Maruyama N., Carp R.I., Kim Y.S., Choi E.K. Peptidylarginine deiminase modulates the physiological roles of enolase via citrullination: links between altered multifunction of enolase and neurodegenerative diseases. Biochem. J. 2012;445:183-92. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
