

Quantitative proteomics of the Neisseria gonorrhoeae cell envelope and membrane vesicles for the discovery of potential therapeutic targets
- PMID: 24607996
- PMCID: PMC4014286
- DOI: 10.1074/mcp.M113.029538
Quantitative proteomics of the Neisseria gonorrhoeae cell envelope and membrane vesicles for the discovery of potential therapeutic targets
Abstract
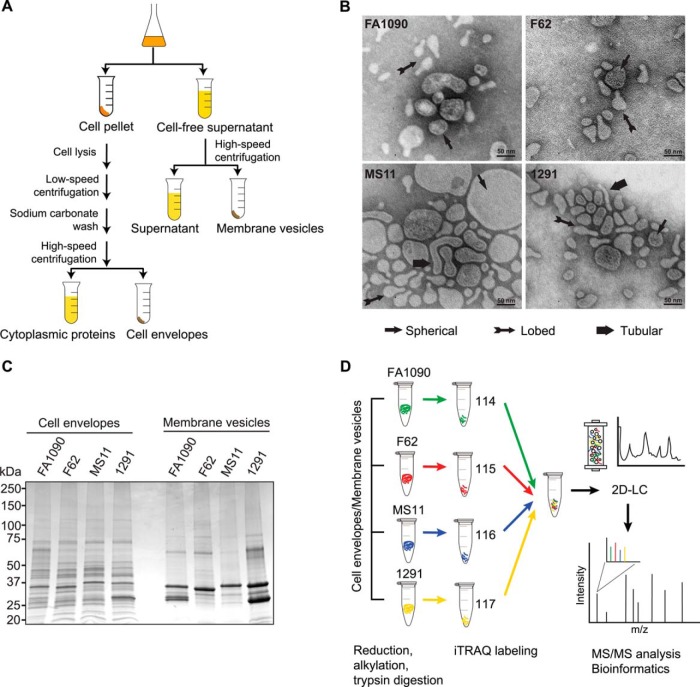
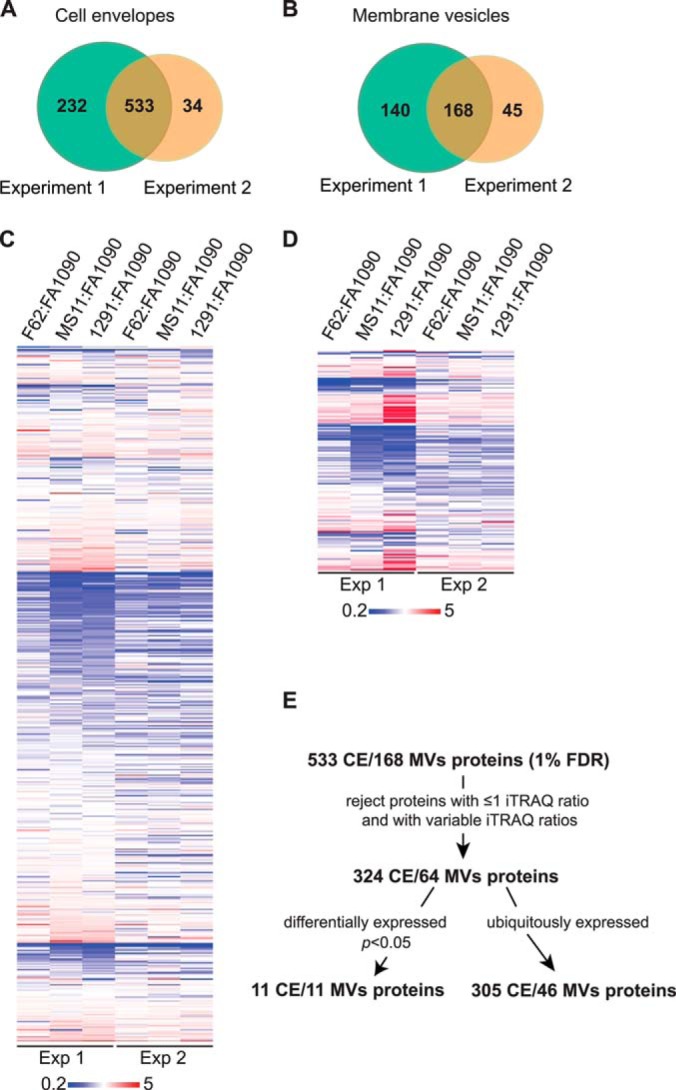
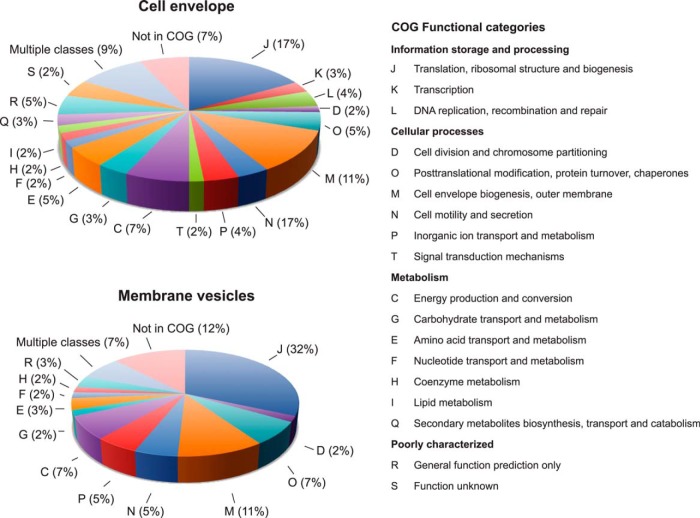
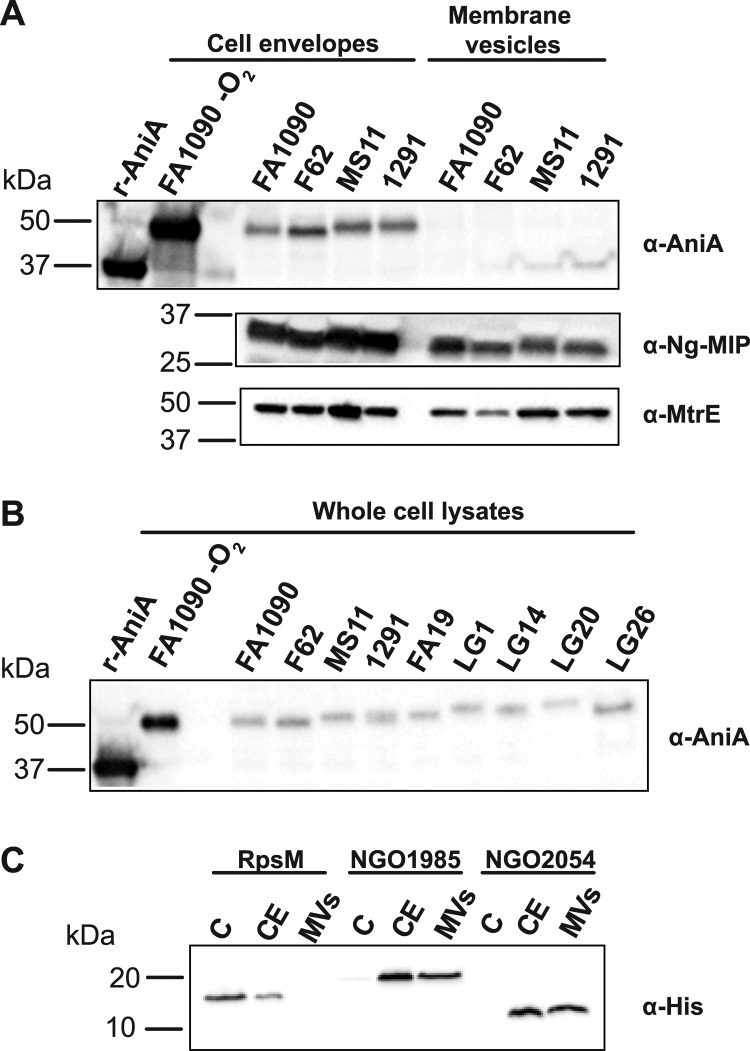
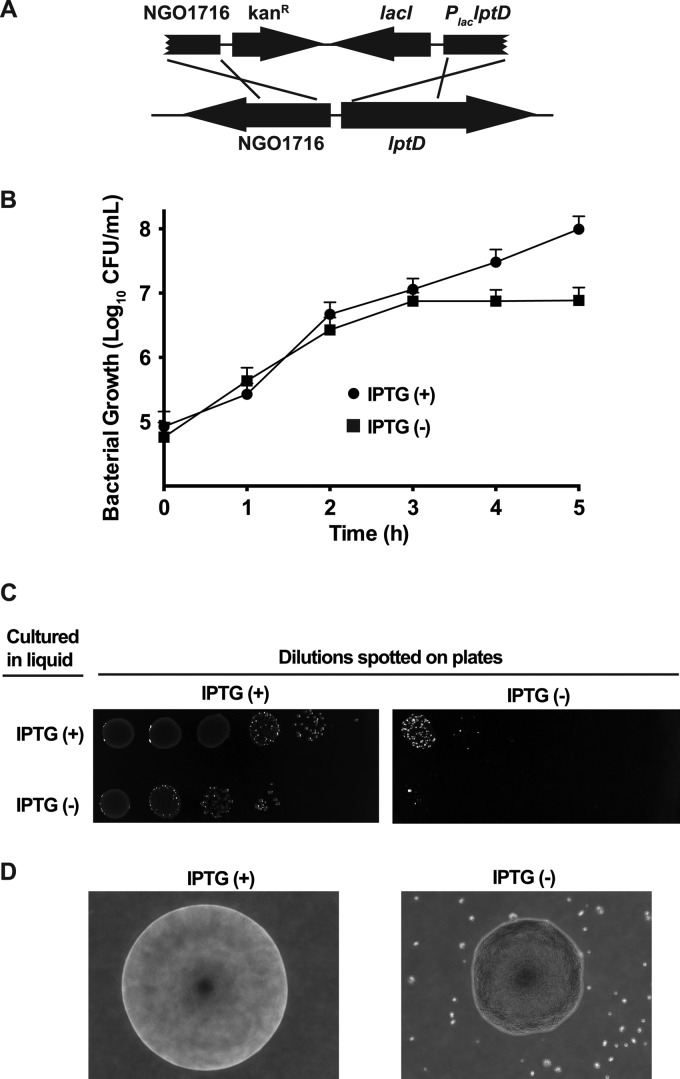
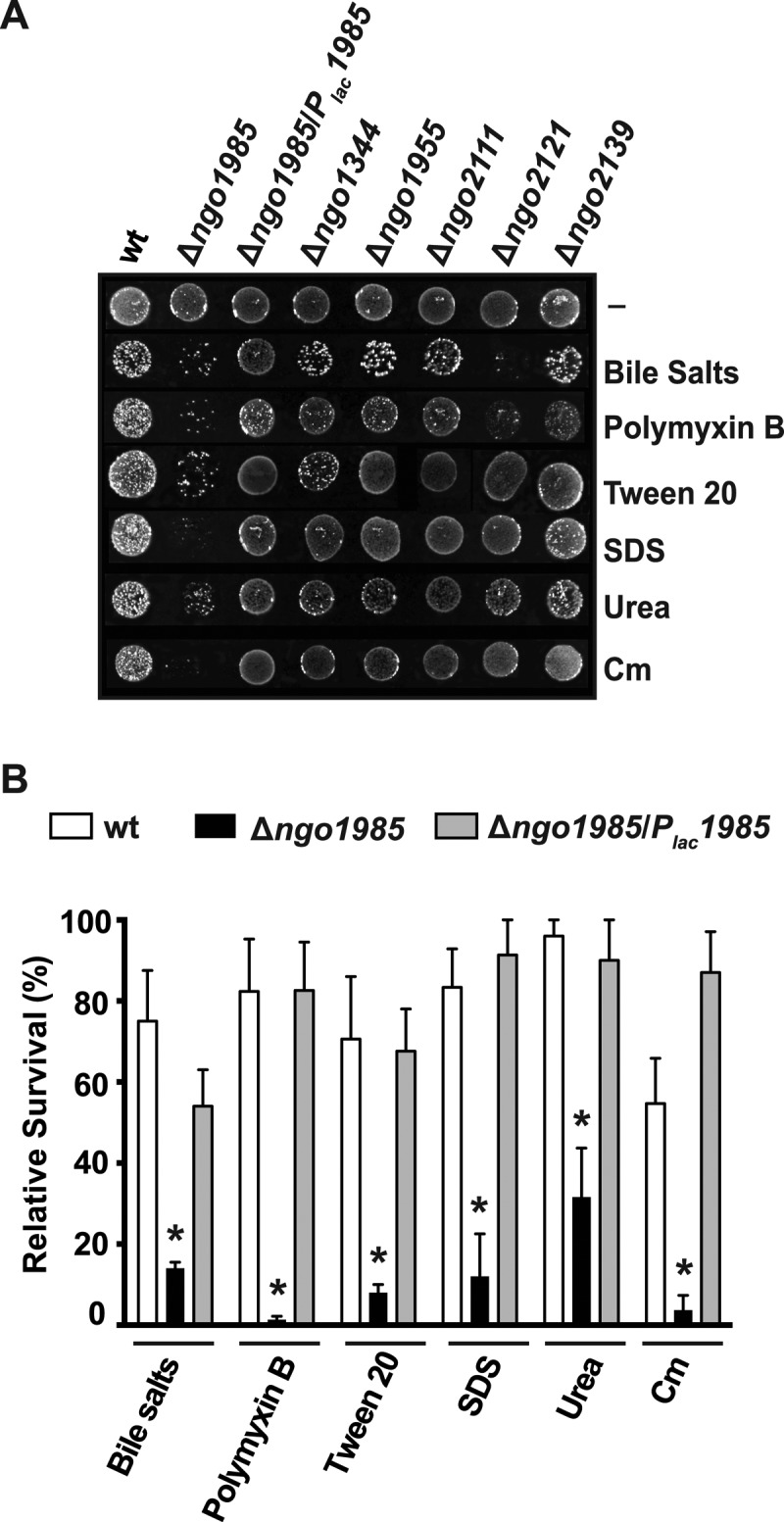
Neisseria gonorrhoeae (GC) is a human-specific pathogen, and the agent of a sexually transmitted disease, gonorrhea. There is a critical need for new approaches to study and treat GC infections because of the growing threat of multidrug-resistant isolates and the lack of a vaccine. Despite the implied role of the GC cell envelope and membrane vesicles in colonization and infection of human tissues and cell lines, comprehensive studies have not been undertaken to elucidate their constituents. Accordingly, in pursuit of novel molecular therapeutic targets, we have applied isobaric tagging for absolute quantification coupled with liquid chromatography and mass spectrometry for proteome quantitative analyses. Mining the proteome of cell envelopes and native membrane vesicles revealed 533 and 168 common proteins, respectively, in analyzed GC strains FA1090, F62, MS11, and 1291. A total of 22 differentially abundant proteins were discovered including previously unknown proteins. Among those proteins that displayed similar abundance in four GC strains, 34 were found in both cell envelopes and membrane vesicles fractions. Focusing on one of them, a homolog of an outer membrane protein LptD, we demonstrated that its depletion caused loss of GC viability. In addition, we selected for initial characterization six predicted outer membrane proteins with unknown function, which were identified as ubiquitous in the cell envelopes derived from examined GC isolates. These studies entitled a construction of deletion mutants and analyses of their resistance to different chemical probes. Loss of NGO1985, in particular, resulted in dramatically decreased GC viability upon treatment with detergents, polymyxin B, and chloramphenicol, suggesting that this protein functions in the maintenance of the cell envelope permeability barrier. Together, these findings underscore the concept that the cell envelope and membrane vesicles contain crucial, yet under-explored determinants of GC physiology, which may represent promising targets for designing new therapeutic interventions.
Figures
References
-
- Lusti-Narasimhan M., Ndowa F. (2011) Emergence and spread of multi-drug resistant Neisseria gonorrhoeae. J. Sex. Med. 8, 253
-
- McNabb S. J., Jajosky R. A., Hall-Baker P. A., Adams D. A., Sharp P., Worshams C., Anderson W. J., Javier A. J., Jones G. J., Nitschke D. A., Rey A., Wodajo M. S. (2008) Summary of notifiable diseases–United States, 2006. MMWR Morb. Mortal. Wkly Rep. 55, 1–92 - PubMed
-
- Workowski K. A., Berman S. (2010) Sexually transmitted diseases treatment guidelines, 2010. MMWR Recomm. Rep. 59, 1–110 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
