

Myc-induced anchorage of the rDNA IGS region to nucleolar matrix modulates growth-stimulated changes in higher-order rDNA architecture
- PMID: 24609384
- PMCID: PMC4027186
- DOI: 10.1093/nar/gku183
Myc-induced anchorage of the rDNA IGS region to nucleolar matrix modulates growth-stimulated changes in higher-order rDNA architecture
Abstract
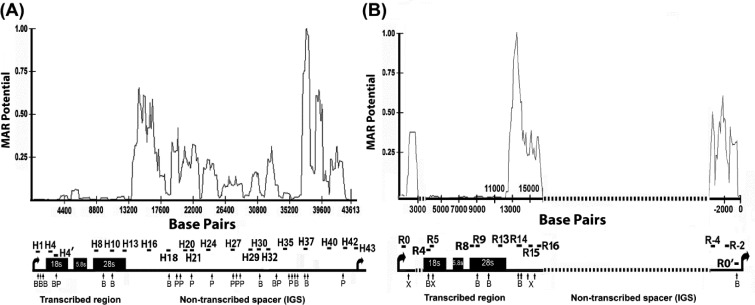
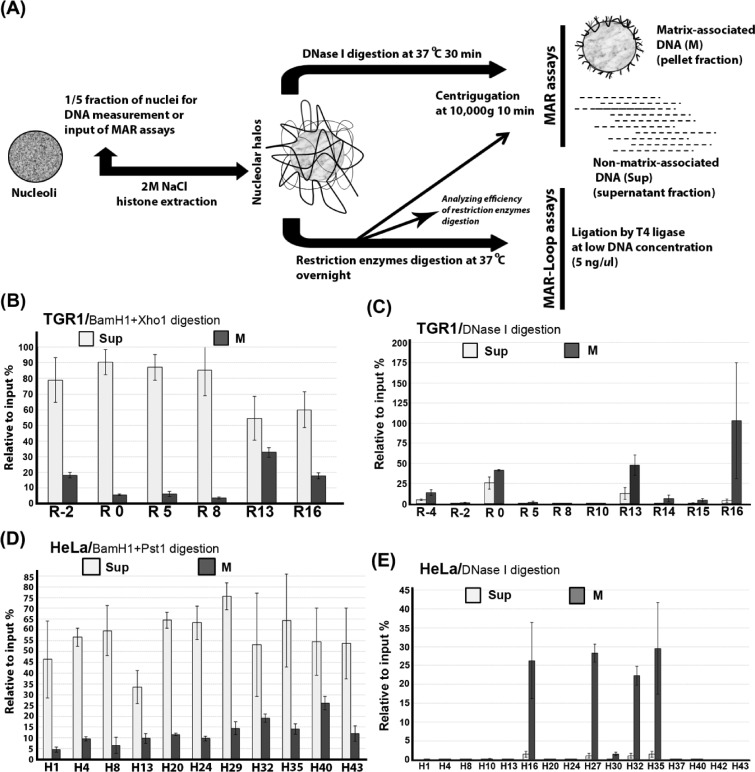
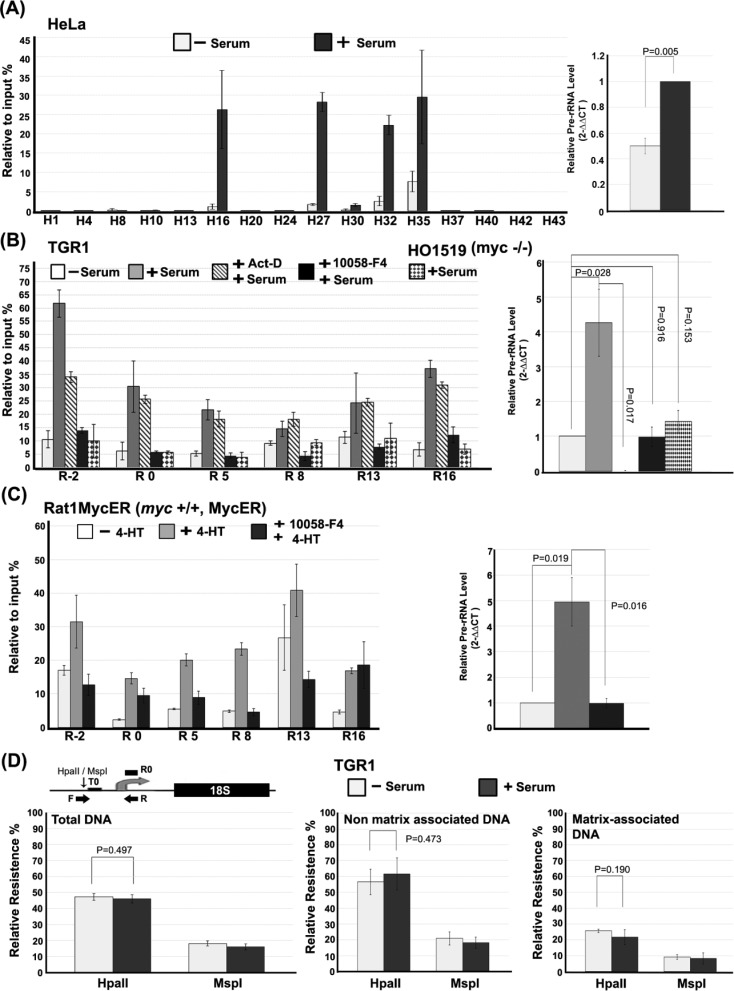
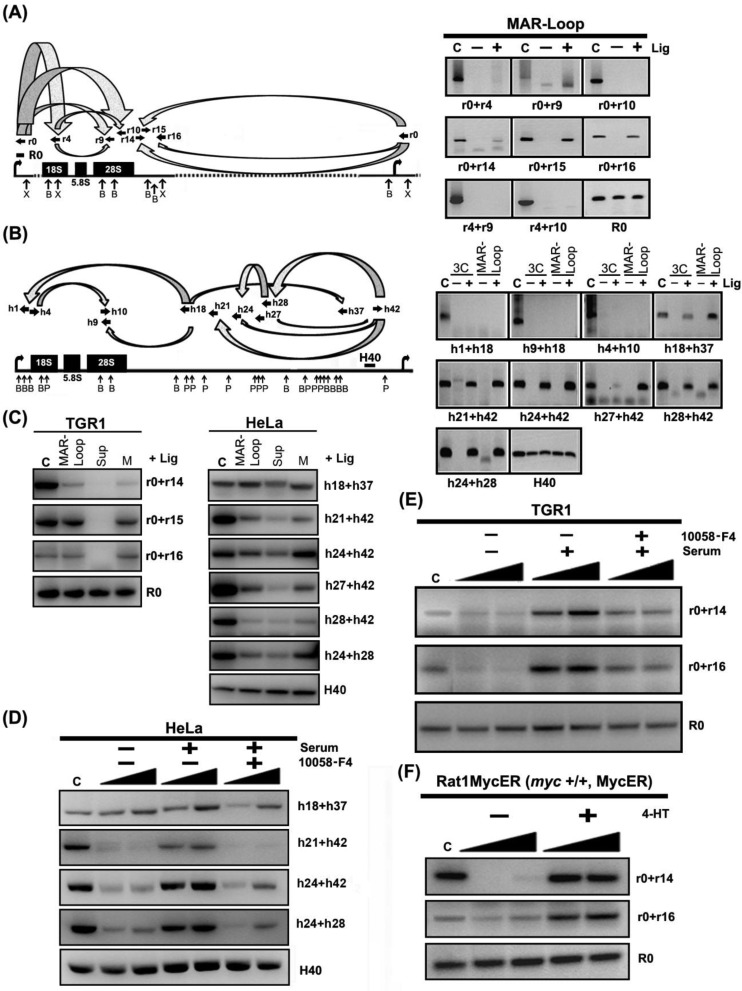
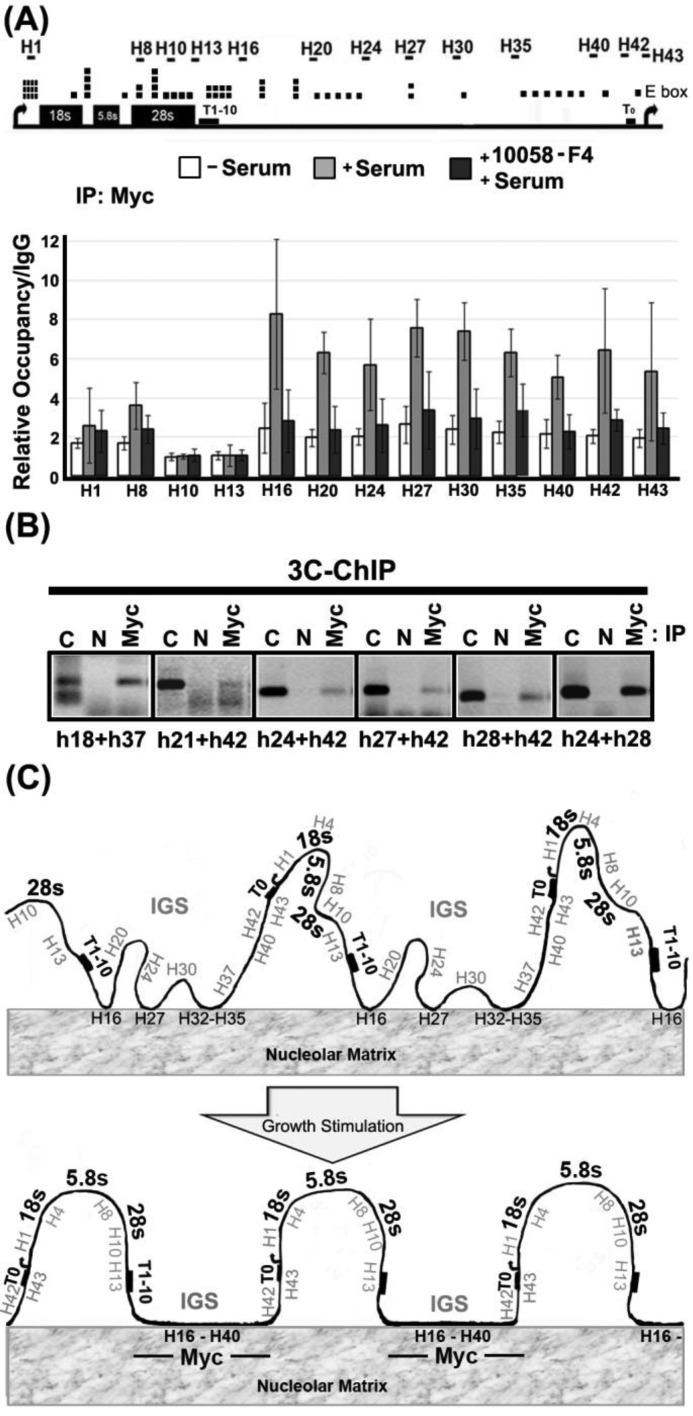
Chromatin domain organization and the compartmentalized distribution of chromosomal regions are essential for packaging of deoxyribonucleic acid (DNA) in the eukaryotic nucleus as well as regulated gene expression. Nucleoli are the most prominent morphological structures of cell nuclei and nucleolar organization is coupled to cell growth. It has been shown that nuclear scaffold/matrix attachment regions often define the base of looped chromosomal domains in vivo and that they are thereby critical for correct chromosome architecture and gene expression. Here, we show regulated organization of mammalian ribosomal ribonucleic acid genes into distinct chromatin loops by tethering to nucleolar matrix via the non-transcribed inter-genic spacer region of the ribosomal DNA (rDNA). The rDNA gene loop structures are induced specifically upon growth stimulation and are dependent on the activity of the c-Myc protein. Matrix-attached rDNA genes are hypomethylated at the promoter and are thus available for transcriptional activation. rDNA genes silenced by methylation are not recruited to the matrix. c-Myc, which has been shown to induce rDNA transcription directly, is physically associated with rDNA gene looping structures and the intergenic spacer sequence in growing cells. Such a role of Myc proteins in gene activation has not been reported previously.
© 2014 The Author(s). Published by Oxford University Press [on behalf of Nucleic Acids Research].
Figures
Similar articles
-
Nucleolar organization, growth control and cancer.Epigenetics. 2010 Apr;5(3):200-5. doi: 10.4161/epi.5.3.11376. Epub 2010 Apr 1. Epigenetics. 2010. PMID: 20305389
-
Loss of nucleolar histone chaperone NPM1 triggers rearrangement of heterochromatin and synergizes with a deficiency in DNA methyltransferase DNMT3A to drive ribosomal DNA transcription.J Biol Chem. 2014 Dec 12;289(50):34601-19. doi: 10.1074/jbc.M114.569244. Epub 2014 Oct 27. J Biol Chem. 2014. PMID: 25349213 Free PMC article.
-
c-Myc induces changes in higher order rDNA structure on stimulation of quiescent cells.Oncogene. 2009 Apr 23;28(16):1833-42. doi: 10.1038/onc.2009.21. Epub 2009 Mar 9. Oncogene. 2009. PMID: 19270725
-
Stress-Induced Evolution of the Nucleolus: The Role of Ribosomal Intergenic Spacer (rIGS) Transcripts.Biomolecules. 2024 Oct 20;14(10):1333. doi: 10.3390/biom14101333. Biomolecules. 2024. PMID: 39456266 Free PMC article. Review.
-
Nucleolar DNA: the host and the guests.Histochem Cell Biol. 2016 Apr;145(4):359-72. doi: 10.1007/s00418-016-1407-x. Epub 2016 Feb 4. Histochem Cell Biol. 2016. PMID: 26847178 Review.
Cited by
-
Acute appendicitis: transcript profiling of blood identifies promising biomarkers and potential underlying processes.BMC Med Genomics. 2016 Jul 15;9(1):40. doi: 10.1186/s12920-016-0200-y. BMC Med Genomics. 2016. PMID: 27417541 Free PMC article.
-
Mapping nucleolus-associated chromatin interactions using nucleolus Hi-C reveals pattern of heterochromatin interactions.Nat Commun. 2023 Jan 21;14(1):350. doi: 10.1038/s41467-023-36021-1. Nat Commun. 2023. PMID: 36681699 Free PMC article.
-
Reproduction of the FC/DFC units in nucleoli.Nucleus. 2016 Apr 25;7(2):203-15. doi: 10.1080/19491034.2016.1157674. Epub 2016 Mar 2. Nucleus. 2016. PMID: 26934002 Free PMC article.
-
Nucleolar RNA polymerase II drives ribosome biogenesis.Nature. 2020 Sep;585(7824):298-302. doi: 10.1038/s41586-020-2497-0. Epub 2020 Jul 15. Nature. 2020. PMID: 32669707 Free PMC article.
-
Nucleolus and chromatin.Histochem Cell Biol. 2018 Sep;150(3):209-225. doi: 10.1007/s00418-018-1696-3. Epub 2018 Jul 25. Histochem Cell Biol. 2018. PMID: 30046888 Free PMC article. Review.
References
-
- Felsenfeld G. Chromatin unfolds. Cell. 1996;86:13–19. - PubMed
-
- Brown K.E. Chromatin folding and gene expression: new tools to reveal the spatial organization of genes. Chromosome Res. 2003;11:423–433. - PubMed
-
- Mehta I.S., Figgitt M., Clements C.S., Kill I.R., Bridger J.M. Alterations to nuclear architecture and genome behavior in senescent cells. Ann. N.Y. Acad. Sci. 2007;1100:250–263. - PubMed
-
- Prieto J.L., McStay B. Nucleolar biogenesis: the first small steps. Biochem. Soc. Trans. 2005;33:1441–1443. - PubMed
-
- Raska I., Shaw P.J., Cmarko D. New insights into nucleolar architecture and activity. Int. Rev. Cytol. 2006;255:177–235. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
