

Anaplasma phagocytophilum surface protein AipA mediates invasion of mammalian host cells
- PMID: 24612118
- PMCID: PMC4115035
- DOI: 10.1111/cmi.12286
Anaplasma phagocytophilum surface protein AipA mediates invasion of mammalian host cells
Abstract
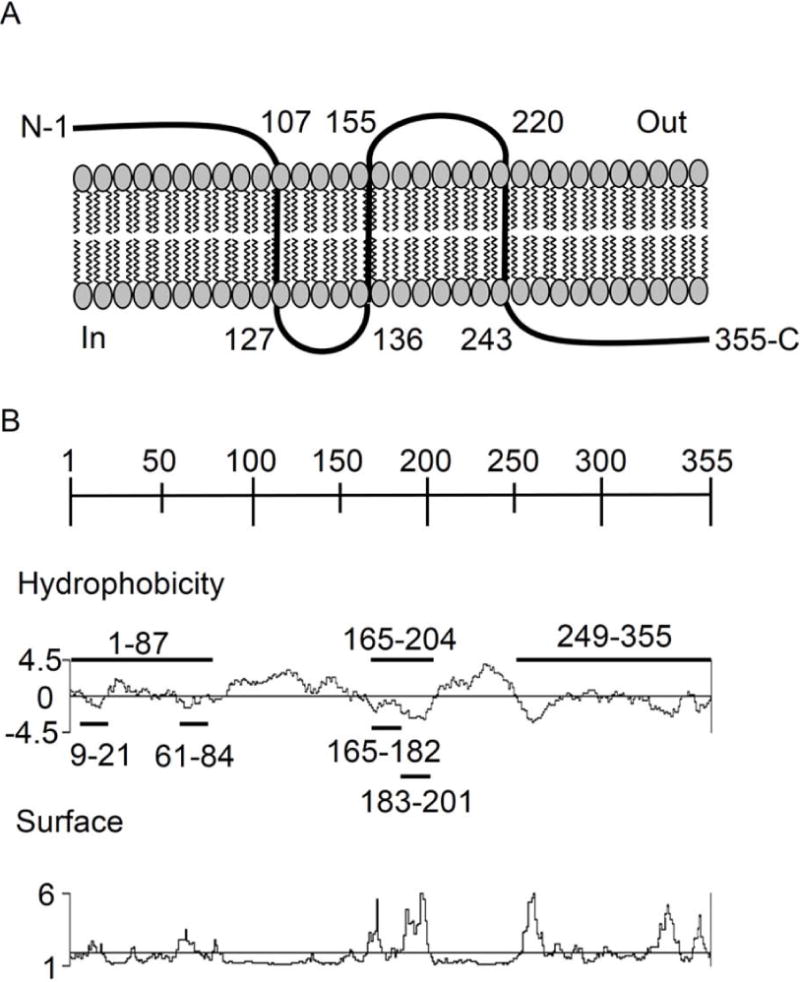
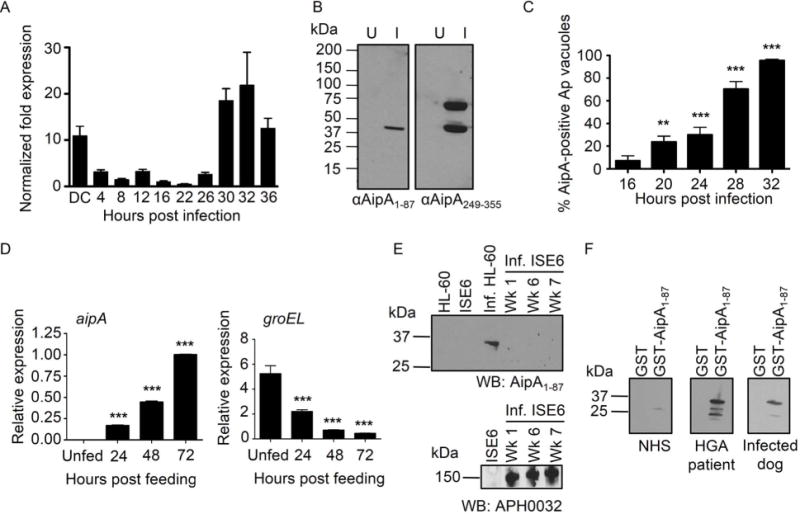
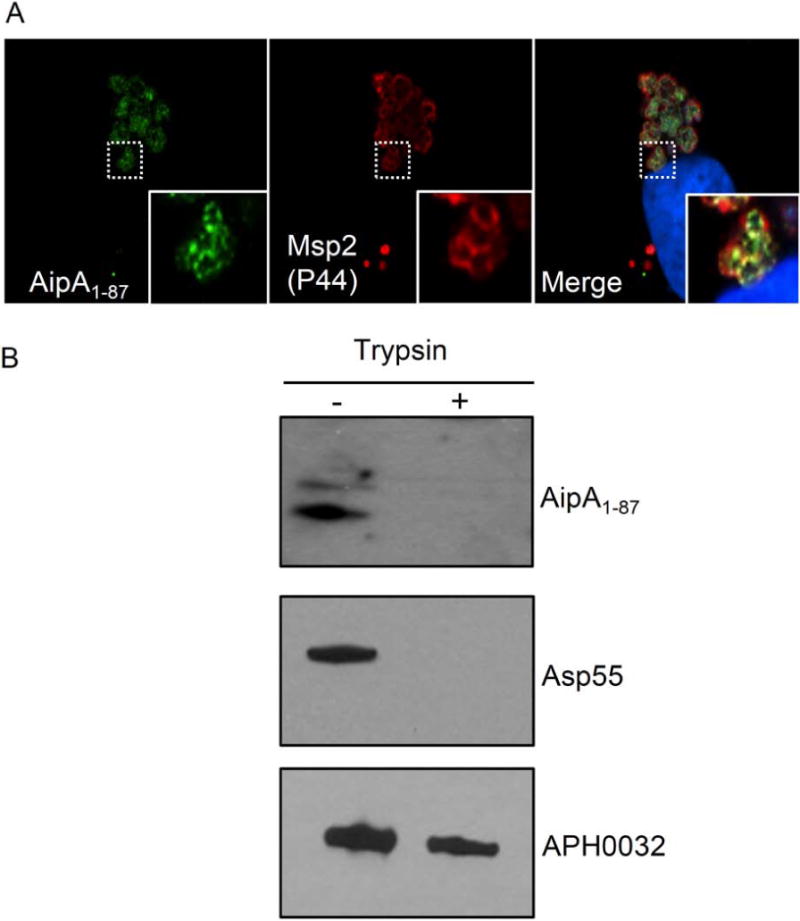
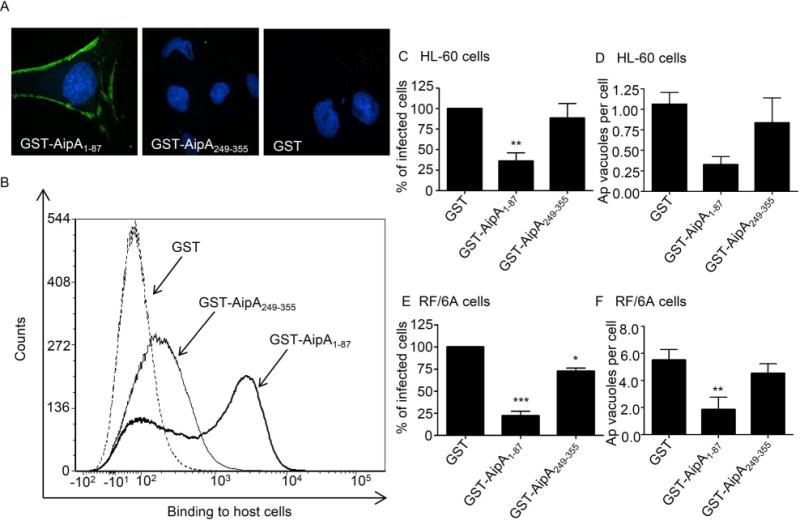
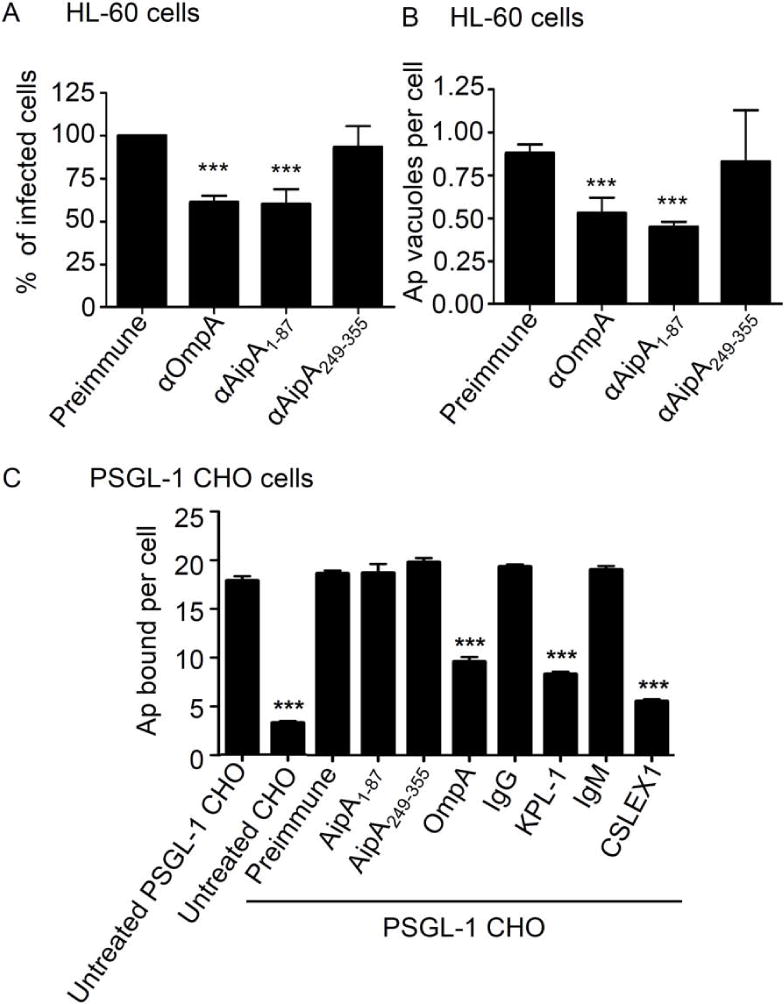
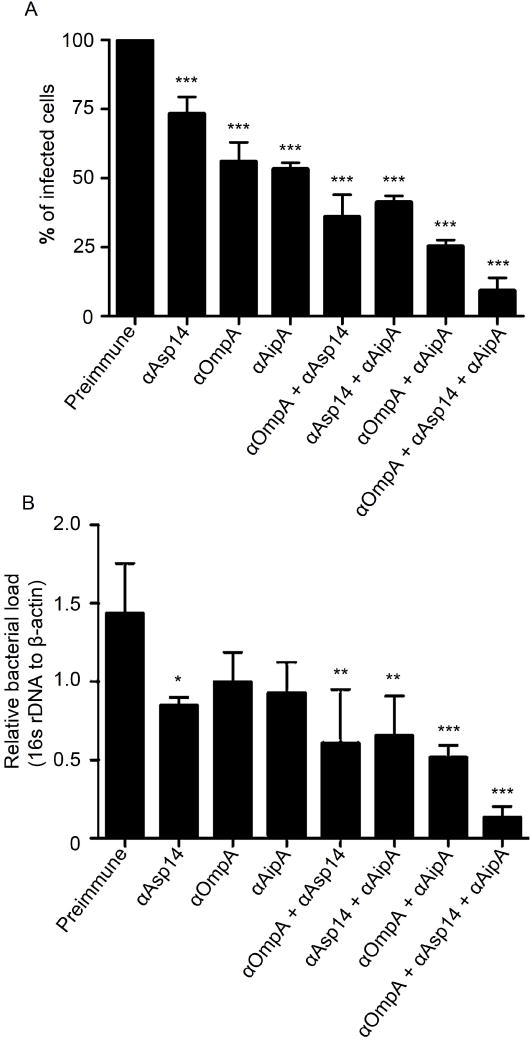
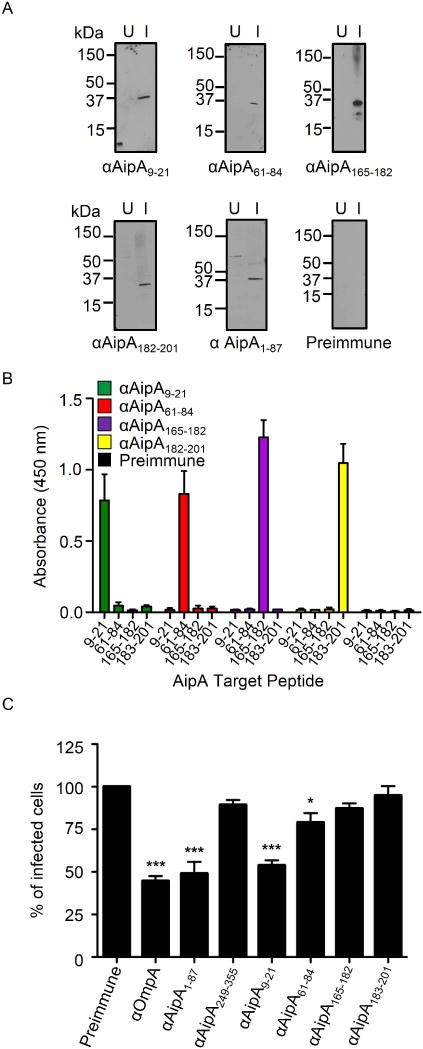
Anaplasma phagocytophilum, which causes granulocytic anaplasmosis in humans and animals, is a tick-transmitted obligate intracellular bacterium that mediates its own uptake into neutrophils and non-phagocytic cells. Invasins of obligate intracellular pathogens are attractive targets for protecting against or curing infection because blocking the internalization step prevents survival of these organisms. The complement of A. phagocytophilum invasins is incompletely defined. Here, we report the significance of a novel A. phagocytophilum invasion protein, AipA. A. phagocytophilum induced aipA expression during transmission feeding of infected ticks on mice. The bacterium upregulated aipA transcription when it transitioned from its non-infectious reticulate cell morphotype to its infectious dense-cored morphotype during infection of HL-60 cells. AipA localized to the bacterial surface and was expressed during in vivo infection. Of the AipA regions predicted to be surface-exposed, only residues 1 to 87 (AipA1-87 ) were found to be essential for host cell invasion. Recombinant AipA1-87 protein bound to and competitively inhibited A. phagocytophilum infection of mammalian cells. Antiserum specific for AipA1-87 , but not other AipA regions, antagonized infection. Additional blocking experiments using peptide-specific antisera narrowed down the AipA invasion domain to residues 9 to 21. An antisera combination targeting AipA1-87 together with two other A. phagocytophilum invasins, OmpA and Asp14, nearly abolished infection of host cells. This study identifies AipA as an A. phagocytophilum surface protein that is critical for infection, demarcates its invasion domain, and establishes a rationale for targeting multiple invasins to protect against granulocytic anaplasmosis.
© 2014 John Wiley & Sons Ltd.
Figures
References
-
- Alhumaidan H, Westley B, Esteva C, Berardi V, Young C, Sweeney J. Transfusion-transmitted anaplasmosis from leukoreduced red blood cells. Transfusion. 2013;53:181–186. - PubMed
-
- Annen K, Friedman K, Eshoa C, Horowitz M, Gottschall J, Straus T. Two cases of transfusion-transmitted Anaplasma phagocytophilum. American journal of clinical pathology. 2012;137:562–565. - PubMed
-
- Brennan MJ, Shahin RD. Pertussis antigens that abrogate bacterial adherence and elicit immunity. American journal of respiratory and critical care medicine. 1996;154:S145–149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
