

Lens β-crystallins: the role of deamidation and related modifications in aging and cataract
- PMID: 24613629
- PMCID: PMC4374893
- DOI: 10.1016/j.pbiomolbio.2014.02.004
Lens β-crystallins: the role of deamidation and related modifications in aging and cataract
Abstract
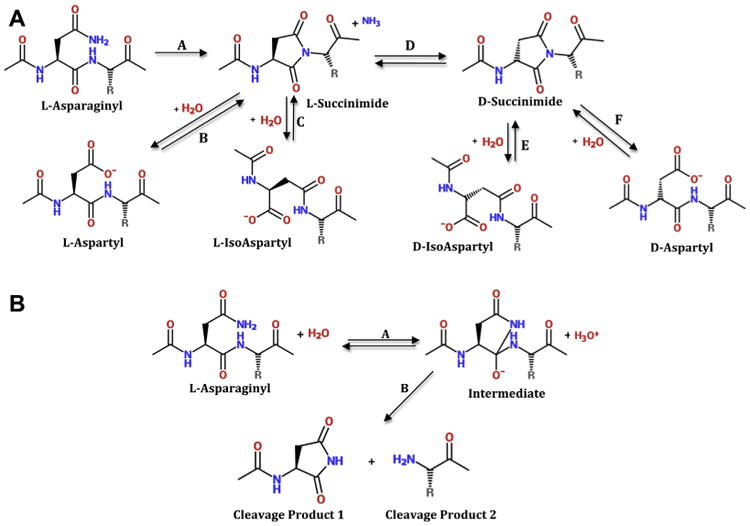
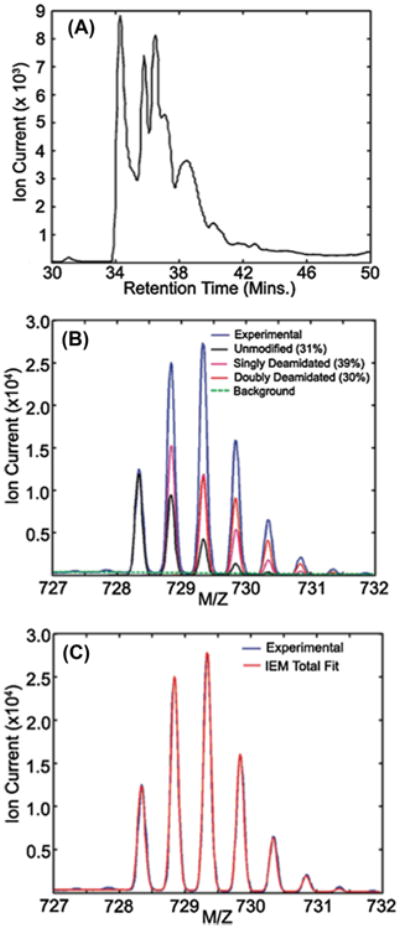
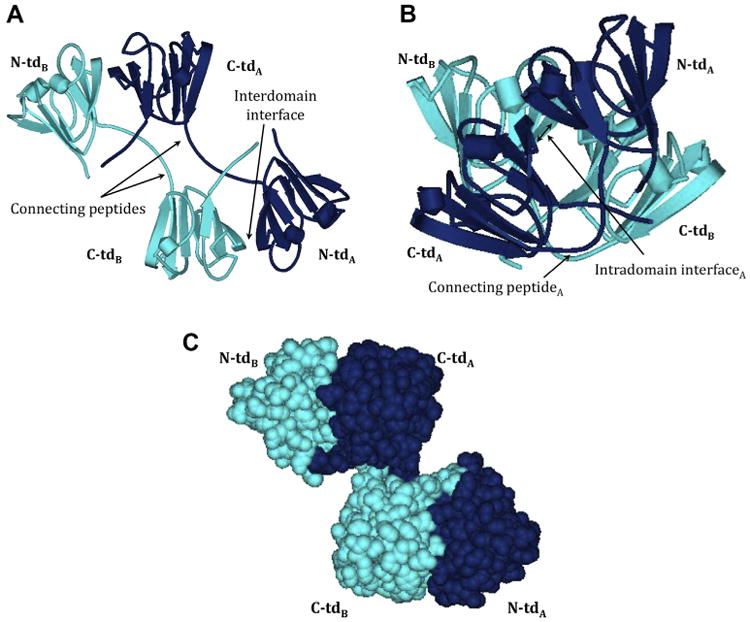
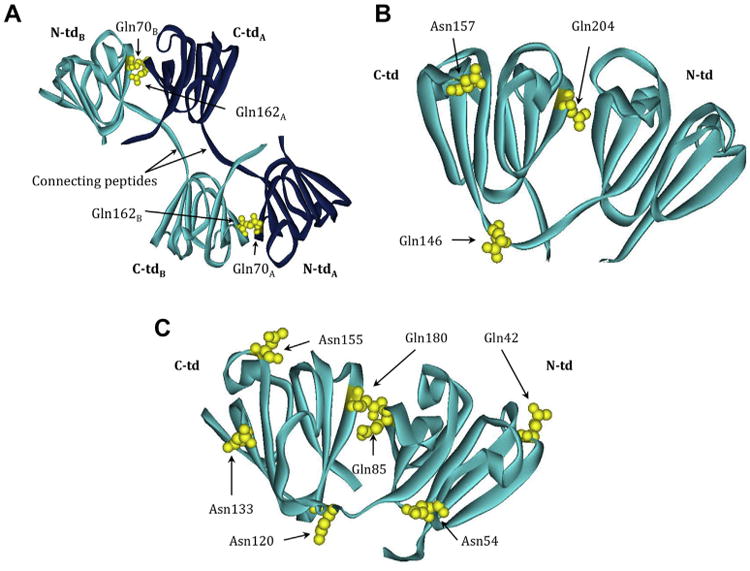
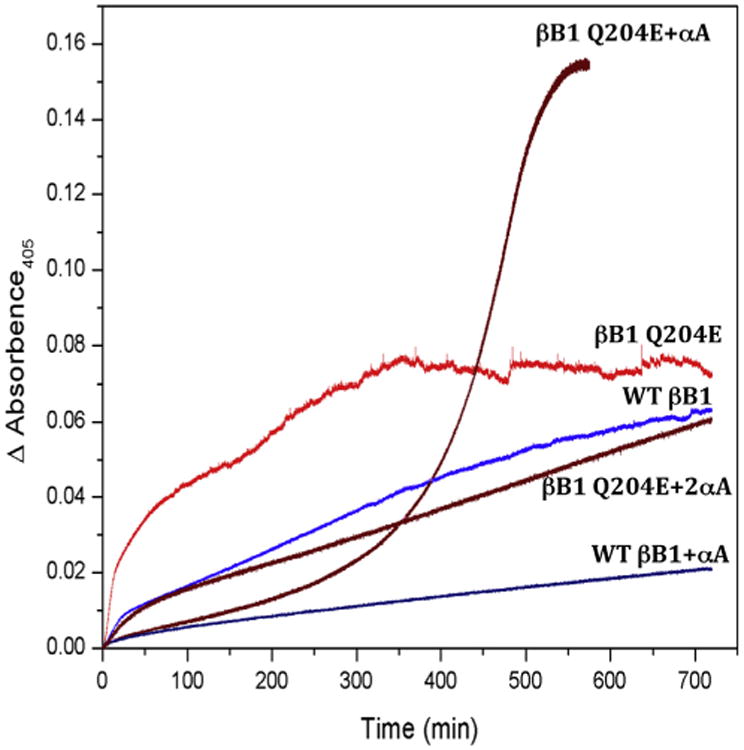
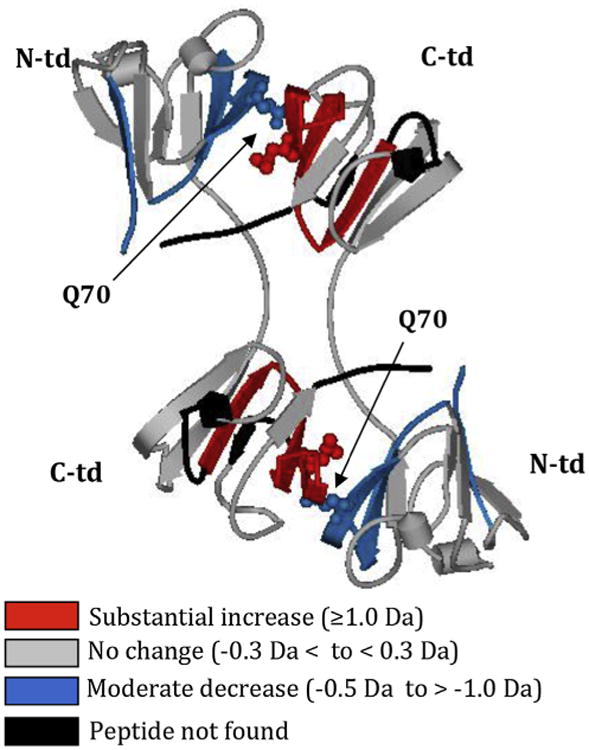
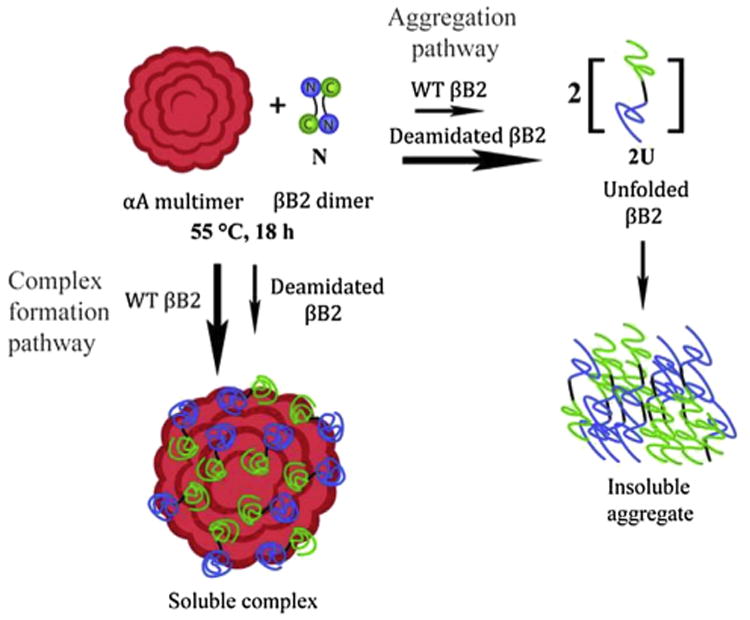
Crystallins are the major proteins in the lens of the eye and function to maintain transparency of the lens. Of the human crystallins, α, β, and γ, the β-crystallins remain the most elusive in their structural significance due to their greater number of subunits and possible oligomer formations. The β-crystallins are also heavily modified during aging. This review focuses on the functional significance of deamidation and the related modifications of racemization and isomerization, the major modifications in β-crystallins of the aged human lens. Elucidating the role of these modifications in cataract formation has been slow, because they are analytically among the most difficult post-translational modifications to study. Recent results suggest that many amides deamidate to similar extent in normal aged and cataractous lenses, while others may undergo greater deamidation in cataract. Mimicking deamidation at critical structural regions induces structural changes that disrupt the stability of the β-crystallins and lead to their aggregation in vitro. Deamidations at the surface disrupt interactions with other crystallins. Additionally, the α-crystallin chaperone is unable to completely prevent deamidated β-crystallins from insolubilization. Therefore, deamidation of β-crystallins may enhance their precipitation and light scattering in vivo contributing to cataract formation. Future experiments are needed to quantify differences in deamidation rates at all Asn and Gln residues within crystallins from aged and cataractous lenses, as well as racemization and isomerization which potentially perturb protein structure greater than deamidation alone. Quantitative data is greatly needed to investigate the importance of these major age-related modifications in cataract formation.
Keywords: Aging; Beta-crystallins; Cataracts; Deamidation; Post-translational modification; Proteomics.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Ajaz MS, Ma Z, Smith DL, Smith JB. Size of human lens beta-crystallin aggregates are distinguished by N-terminal truncation of betaB1. J Biol Chem. 1997;272:11250–11255. - PubMed
-
- Aquilina JA, Benesch JL, Ding LL, Yaron O, Horwitz J, Robinson CV. Subunit exchange of polydisperse proteins: mass spectrometry reveals consequences of alphaA-crystallin truncation. J Biol Chem. 2005;280:14485–14491. - PubMed
-
- Ball LE, Garland DL, Crouch RK, Schey KL. Post-translational modifications of aquaporin 0 (AQP0) in the normal human lens: spatial and temporal occurrence. Biochemistry. 2004;43:9856–9865. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
