

Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities
- PMID: 24617569
- PMCID: PMC4257071
- DOI: 10.1111/1574-6976.12072
Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities
Abstract
Bacteria-phage coevolution, the reciprocal evolution between bacterial hosts and the phages that infect them, is an important driver of ecological and evolutionary processes in microbial communities. There is growing evidence from both laboratory and natural populations that coevolution can maintain phenotypic and genetic diversity, increase the rate of bacterial and phage evolution and divergence, affect community structure, and shape the evolution of ecologically relevant bacterial traits. Although the study of bacteria-phage coevolution is still in its infancy, with open questions regarding the specificity of the interaction, the gene networks of coevolving partners, and the relative importance of the coevolving interaction in complex communities and environments, there have recently been major advancements in the field. In this review, we sum up our current understanding of bacteria-phage coevolution both in the laboratory and in nature, discuss recent findings on both the coevolutionary process itself and the impact of coevolution on bacterial phenotype, diversity and interactions with other species (particularly their eukaryotic hosts), and outline future directions for the field.
Keywords: antagonistic; bacteriophage; host-parasite; infection; resistance; species interaction.
© 2014 The Authors. FEMS Microbiology Reviews published by John Wiley & Sons Ltd on behalf of Federation of European Microbiological Societies.
Figures
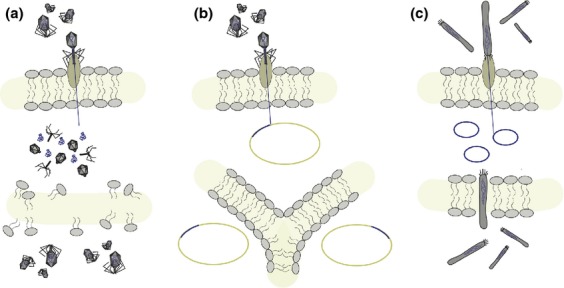
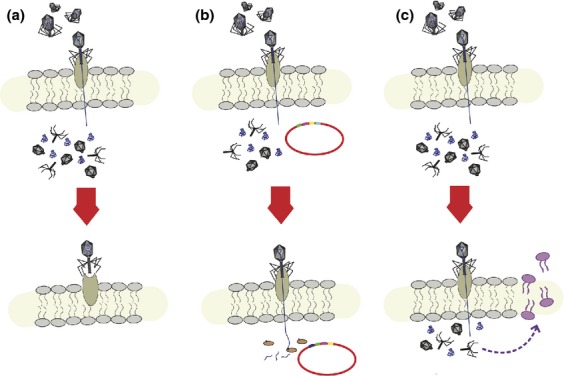
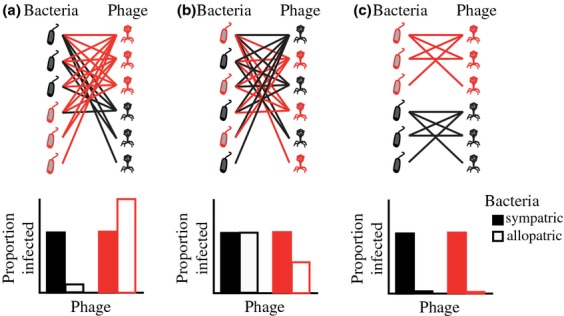
References
-
- Abedon ST. Phages. In: ST Abedon, P Hyman., editors. Bacteriophages in Health and Disease. Vol. 24. MA: CABI, Cambridge; 2012. pp. 1–5.
-
- Allen B, Willner D, Oechel WC. Lipson D. Top-down control of microbial activity and biomass in an Arctic soil ecosystem. Environ Microbiol. 2010;12:642–648. - PubMed
-
- Andersson AF. Banfield JF. Virus population dynamics and acquired virus resistance in natural microbial communities. Science. 2008;320:1047–1050. - PubMed
-
- Avrani S, Wurtzel O, Sharon I, Sorek R. Lindell D. Genomic island variability facilitates Prochlorococcus-virus coexistence. Nature. 2011;474:604–608. - PubMed
-
- Bailey MJ, Lilley AK, Thompson IP, Rainey PB. Ellis RJ. Site directed chromosomal marking of a fluorescent pseudomonad isolated from the phytosphere of sugar beet; Stability and potential for marker gene transfer. Mol Ecol. 1995;4:755–763. - PubMed
