

The evolution of the vertebrate cerebellum: absence of a proliferative external granule layer in a non-teleost ray-finned fish
- PMID: 24617988
- PMCID: PMC5042320
- DOI: 10.1111/ede.12067
The evolution of the vertebrate cerebellum: absence of a proliferative external granule layer in a non-teleost ray-finned fish
Abstract
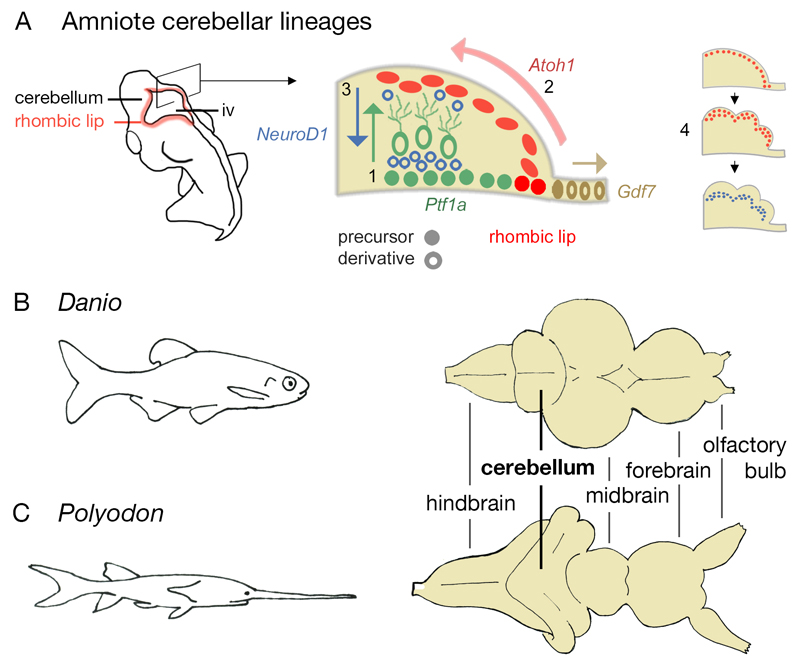
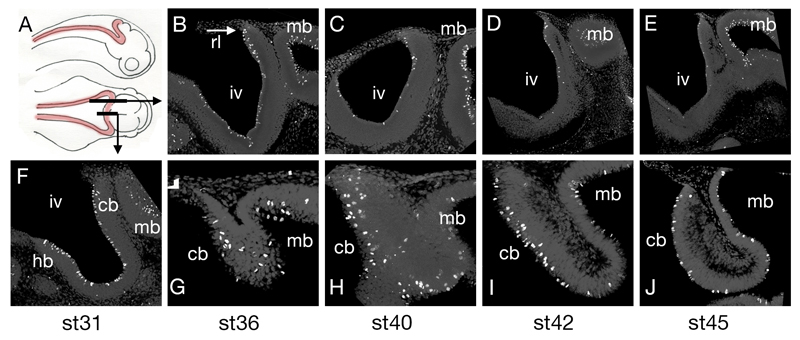
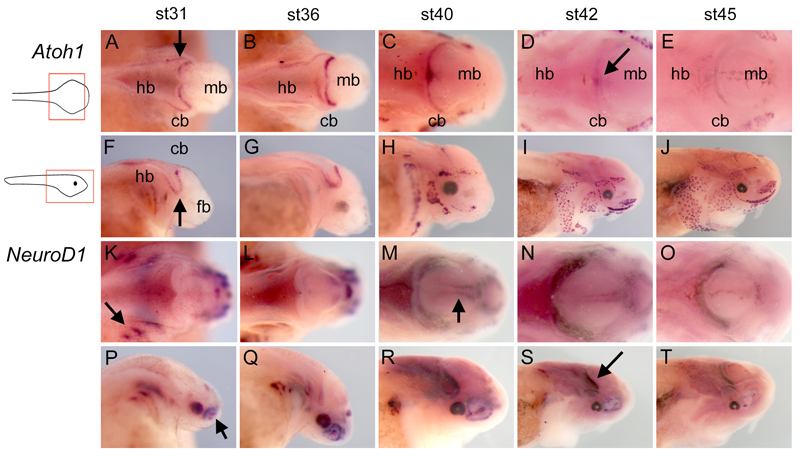
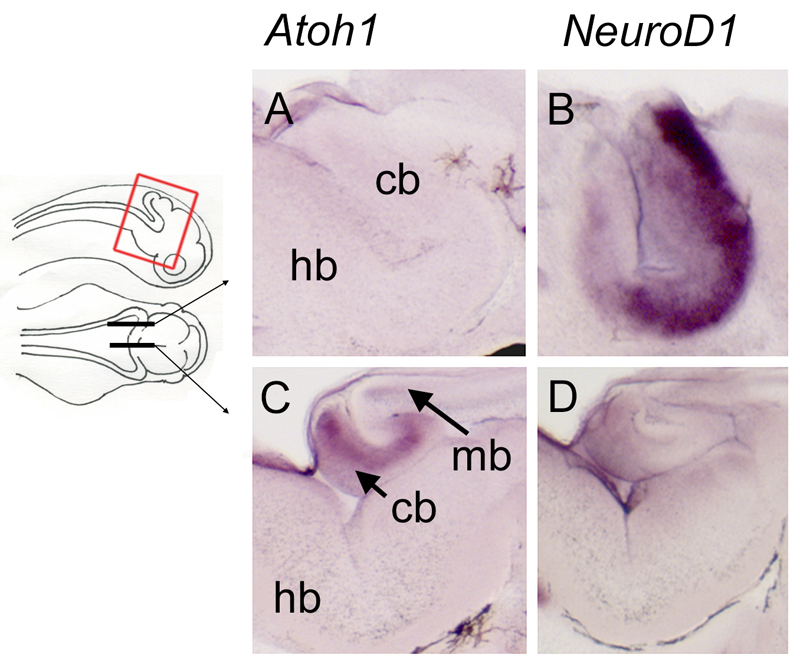
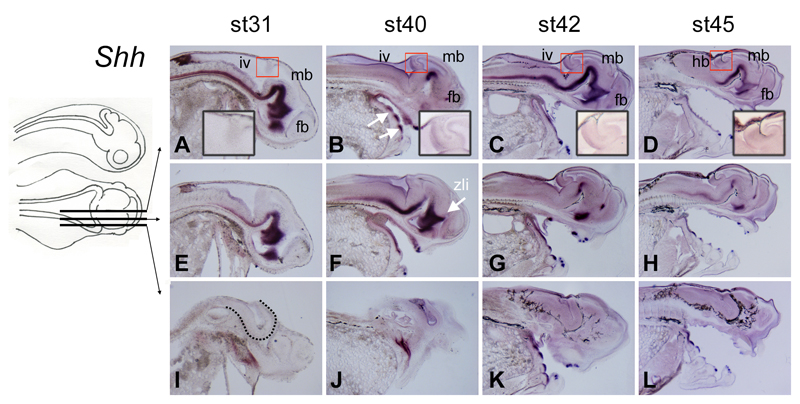
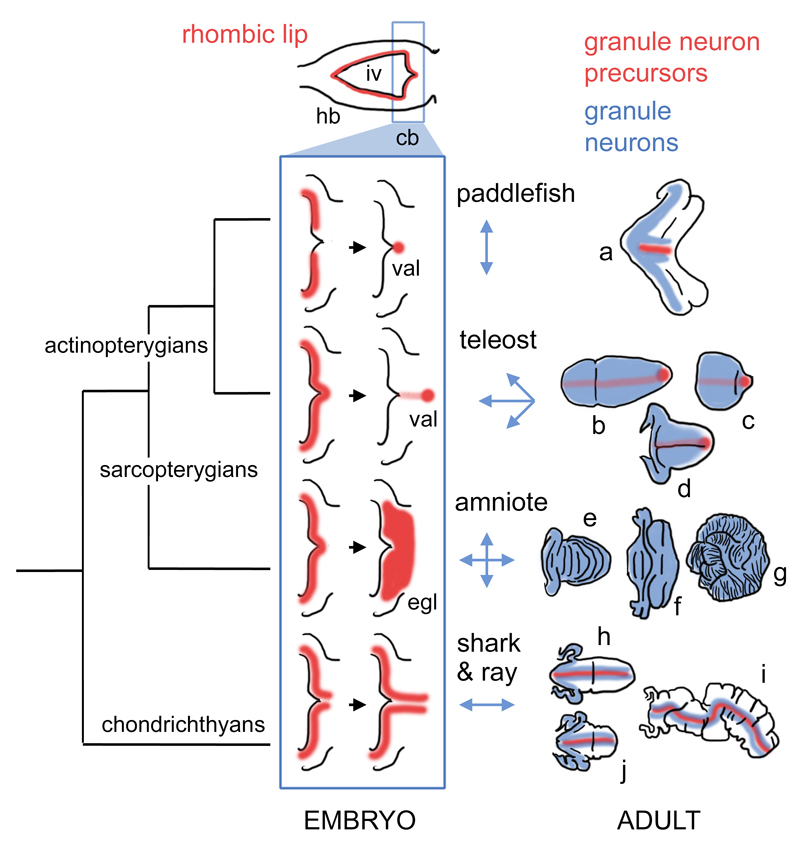
The cerebellum represents one of the most morphologically variable structures in the vertebrate brain. To shed light on its evolutionary history, we have examined the molecular anatomy and proliferation of the developing cerebellum of the North American paddlefish, Polyodon spathula. Absence of an external proliferative cerebellar layer and the restriction of Atonal1 expression to the rhombic lip and valvular primordium demonstrate that transit amplification in a cerebellar external germinal layer, a prominent feature of amniote cerebellum development, is absent in paddlefish. Furthermore, expression of Sonic hedgehog, which drives secondary proliferation in the mouse cerebellum, is absent from the paddlefish cerebellum. These data are consistent with what has been observed in zebrafish and suggest that the transit amplification seen in the amniote cerebellum was either lost very early in the ray-finned fish lineage or evolved in the lobe-finned fish lineage. We also suggest that the Atoh1-positive proliferative valvular primordium may represent a synapomorphy (shared derived character) of ray-finned fishes. The topology of valvular primordium development in paddlefish differs significantly from that of zebrafish and correlates with the adult cerebellar form. The distribution of proliferative granule cell precursors in different vertebrate taxa is thus the likely determining factor in cerebellar morphological diversity.
© 2014 Wiley Periodicals, Inc.
Figures
References
-
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005;21(9):2104–5. - PubMed
-
- Alder J, Cho NK, Hatten ME. Embryonic precursor cells from the rhombic lip are specified to a cerebellar granule neuron identity. Neuron. 1996;17(3):389–99. - PubMed
-
- Bemis WE, Grande L. Early development of the acitnopterygian head. 1. External development and staging of the paddlefish Polydon spatula. Journal of Morphology. 1992;213:47–83. - PubMed
-
- Ben-Arie N, Bellen HJ, Armstrong DL, McCall AE, Gordadze PR, Guo Q, Matzuk MM, Zoghbi HY. Math1 is essential for genesis of cerebellar granule neurons. Nature. 1997;390(6656):169–72. - PubMed
-
- Braitenberg V, Atwood RP. Morphological observations on the cerebellar cortex. J Comp Neurol. 1958;109:1–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/F00818X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F020570/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0901899/MRC_/Medical Research Council/United Kingdom
- BB/I021507/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
