

From cholesterogenesis to steroidogenesis: role of riboflavin and flavoenzymes in the biosynthesis of vitamin D
- PMID: 24618756
- PMCID: PMC3951797
- DOI: 10.3945/an.113.005181
From cholesterogenesis to steroidogenesis: role of riboflavin and flavoenzymes in the biosynthesis of vitamin D
Abstract
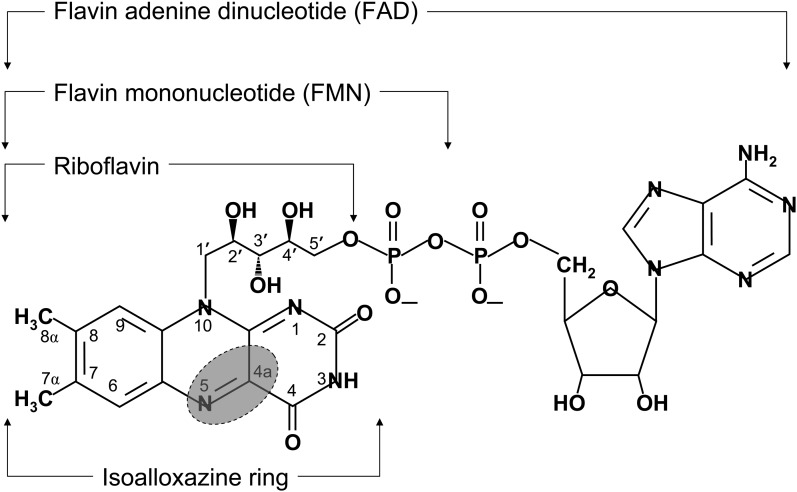
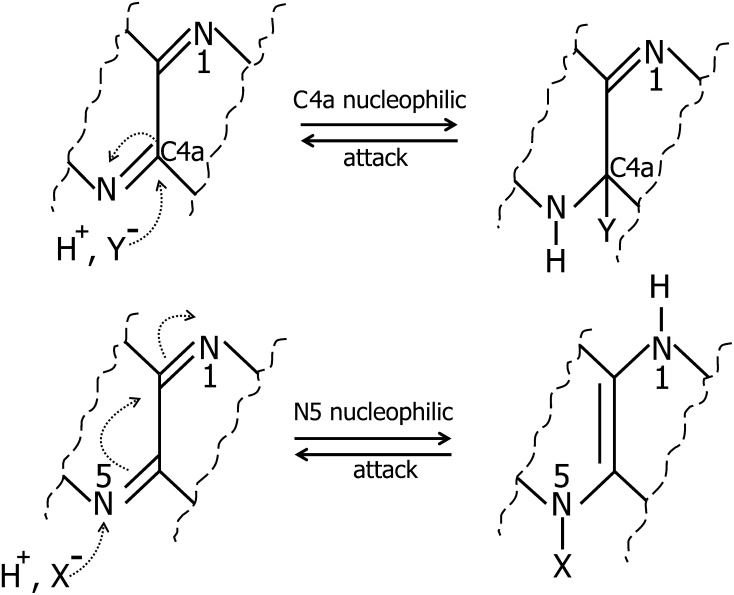
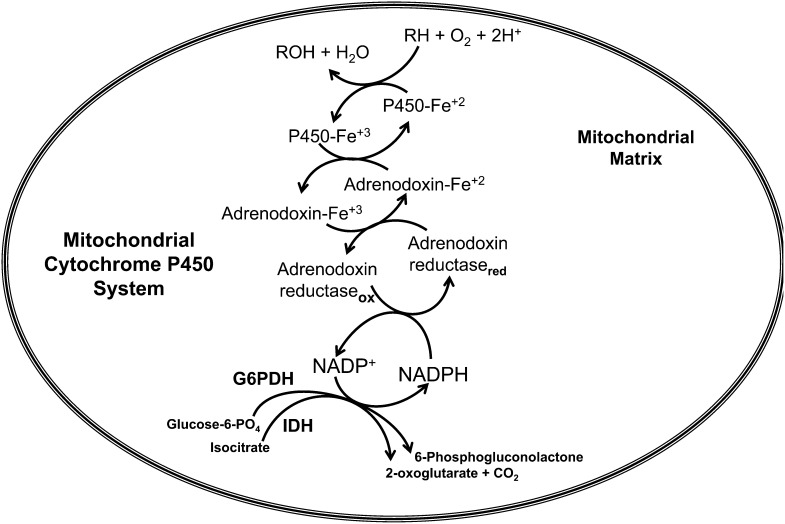
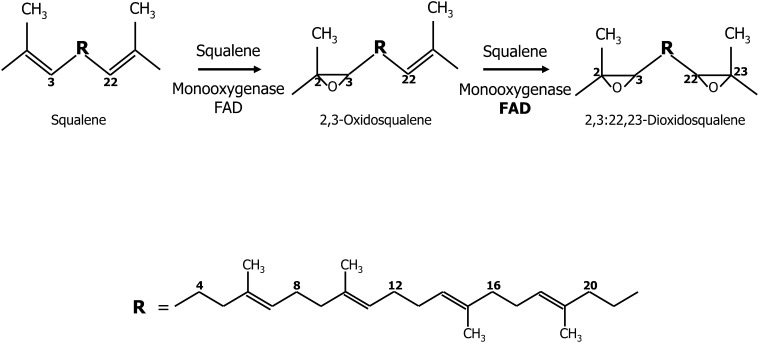
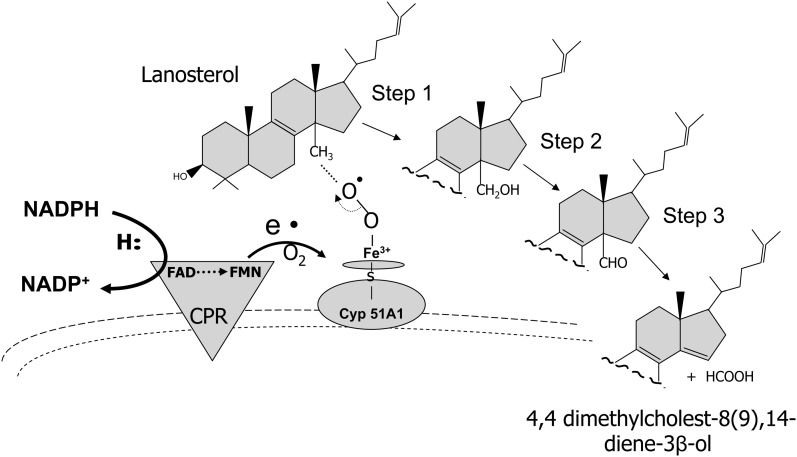
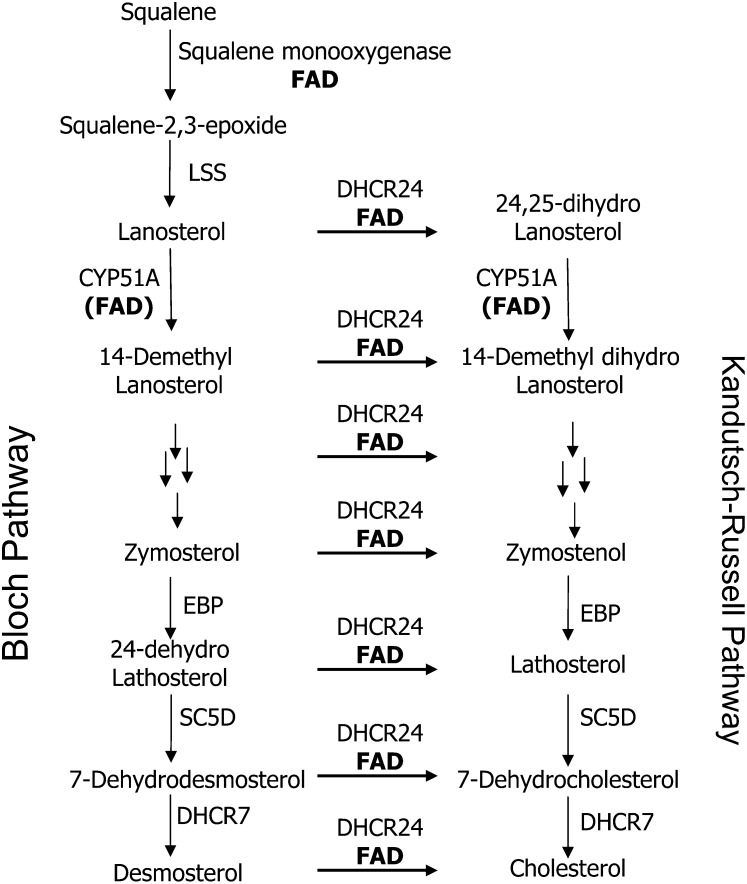
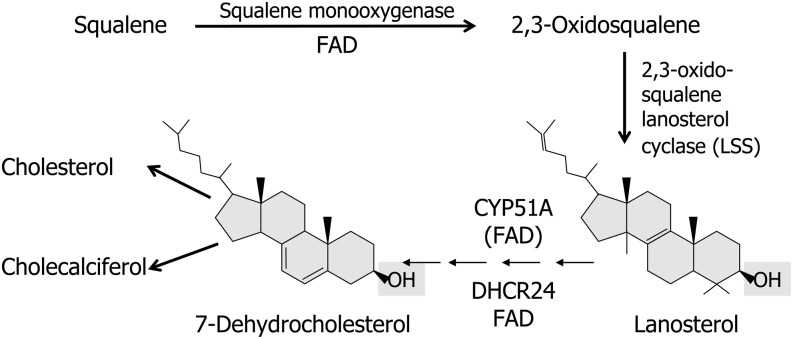
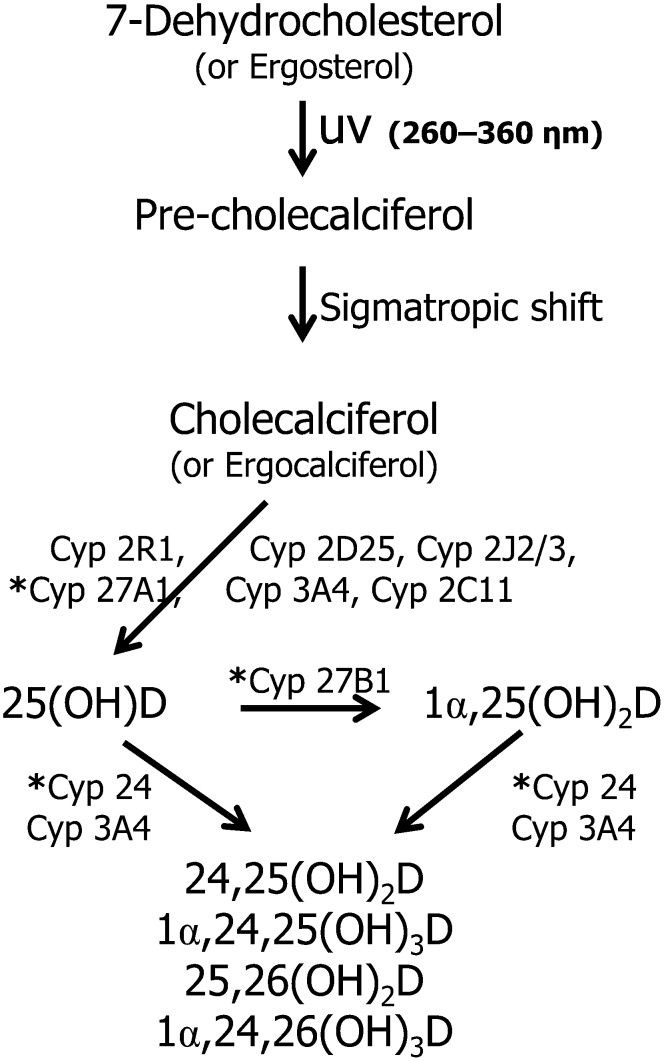
Flavin-dependent monooxygenases and oxidoreductases are located at critical branch points in the biosynthesis and metabolism of cholesterol and vitamin D. These flavoproteins function as obligatory intermediates that accept 2 electrons from NAD(P)H with subsequent 1-electron transfers to a variety of cytochrome P450 (CYP) heme proteins within the mitochondria matrix (type I) and the (microsomal) endoplasmic reticulum (type II). The mode of electron transfer in these systems differs slightly in the number and form of the flavin prosthetic moiety. In the type I mitochondrial system, FAD-adrenodoxin reductase interfaces with adrenodoxin before electron transfer to CYP heme proteins. In the microsomal type II system, a diflavin (FAD/FMN)-dependent cytochrome P450 oxidoreductase [NAD(P)H-cytochrome P450 reductase (CPR)] donates electrons to a multitude of heme oxygenases. Both flavoenzyme complexes exhibit a commonality of function with all CYP enzymes and are crucial for maintaining a balance of cholesterol and vitamin D metabolites. Deficits in riboflavin availability, imbalances in the intracellular ratio of FAD to FMN, and mutations that affect flavin binding domains and/or interactions with client proteins result in marked structural alterations within the skeletal and central nervous systems similar to those of disorders (inborn errors) in the biosynthetic pathways that lead to cholesterol, steroid hormones, and vitamin D and their metabolites. Studies of riboflavin deficiency during embryonic development demonstrate congenital malformations similar to those associated with genetic alterations of the flavoenzymes in these pathways. Overall, a deeper understanding of the role of riboflavin in these pathways may prove essential to targeted therapeutic designs aimed at cholesterol and vitamin D metabolism.
Conflict of interest statement
Author disclosures: J. T. Pinto and A. J. L. Cooper, no conflicts of interest.
Figures
References
-
- Pinto JT, Rivlin RS. Riboflavin In:Zempleni J, Suttie JW, Gregory JF, Stover PJ. editors. 5th ed. New York: Taylor and Francis; 2013. p. 191–265
-
- Nakagawa H, Asano A, Sato R. Ascorbate-synthesizing system in rat liver microsomes. II. A peptide-bound flavin as the prosthetic group of L-gulono-gamma-lactone oxidase. J Biochem. 1975;77:221–32 - PubMed
-
- Kiuchi K, Nishikimi M, Yagi K. L-gulonolactone oxidase activity and vitamin C status in riboflavin-deficient rats. Biochim Biophys Acta. 1980;630:330–7 - PubMed
-
- Mushtaq S, Su H, Hill MH, Powers HJ. Erythrocyte pyridoxamine phosphate oxidase activity: a potential biomarker of riboflavin status? Am J Clin Nutr. 2009;90:1151–9 - PubMed
-
- Adelekan DA, Adekile AD, Thurnham DI. Dependence of pyridoxine metabolism on riboflavin status in sickle cell patients. Am J Clin Nutr. 1987;46:86–90 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
