

doi: 10.1534/genetics.114.163824.
Epub 2014 Mar 12.
Degradation of specific nuclear proteins occurs in the cytoplasm in Saccharomyces cerevisiae
Affiliations
- PMID: 24621829
- PMCID: PMC4012479
- DOI: 10.1534/genetics.114.163824
Item in Clipboard
Degradation of specific nuclear proteins occurs in the cytoplasm in Saccharomyces cerevisiae
Genetics.
2014 May.
Abstract
The ubiquitin/proteasome system has been characterized extensively, although the site of nuclear substrate turnover has not been established definitively. We report here that two well-characterized nuclear proteins are stabilized in nuclear export mutants in Saccharomyces cerevisiae. The requirement for nuclear export defines a new regulatory step in intracellular proteolysis.
Keywords: degradation; export; proteasome; substrate; ubiquitin.
Figures
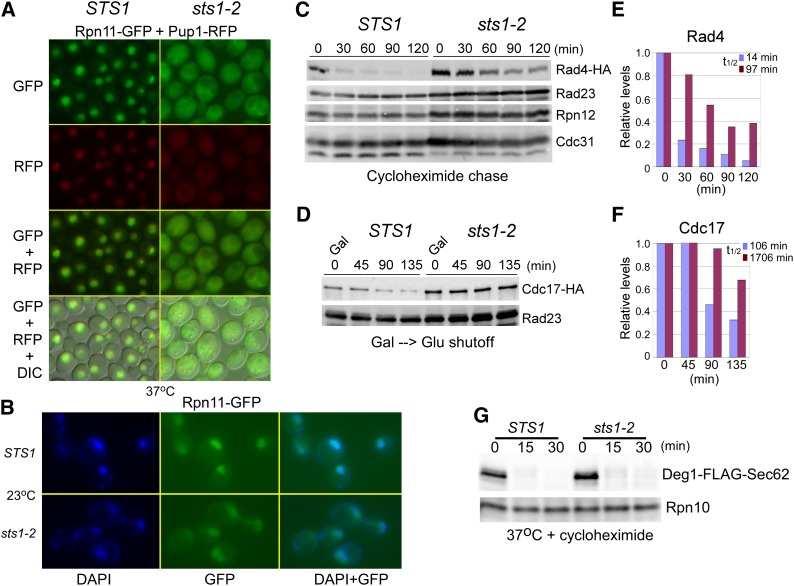
Nuclear substrates are stabilized in sts1-2. (A) The localization of proteasome subunits Rpn11–GFP (19S) and Pup1–RFP (20S) was investigated in STS1 and sts1-2. Both subunits were nuclear localized in STS1, but were mislocalized in sts1-2. A merged image including DIC is also shown. (B) The colocalization of proteasome subunit Rpn11–GFP with the nucleus was confirmed by staining STS1 and sts1-2 cells with DAPI. (C and D) We compared the stability of HA-tagged nuclear proteasome substrates (Rad4–HA; Cdc17–HA) in STS1 and sts1-2. Cultures expressing Rad4–HA were transferred from 23° to 37°, and cycloheximide was added. Protein extracts were prepared at the times indicated and analyzed by immunoblotting. The filter was probed with antibodies against HA, Rad23, Rpn12, and Cdc31. Cdc17–HA was expressed from the regulated GAL1 promoter at 23°, expression was inhibited after transfer to glucose medium at 37°, and samples were examined at the times indicated. The blot was treated with antibodies against HA and Rad23. The results in C and D were quantified by densitometry and values representing one of three independent studies are shown (E and F). Nuclear substrate levels were standardized to the level of the loading control, and the 0-min value was set to 1.0. (G) The stability of the cytosolic proteasome substrate Deg1–FLAG–Sec62 was examined in STS1 and sts1-2 (at 37°), after the addition of cycloheximide.
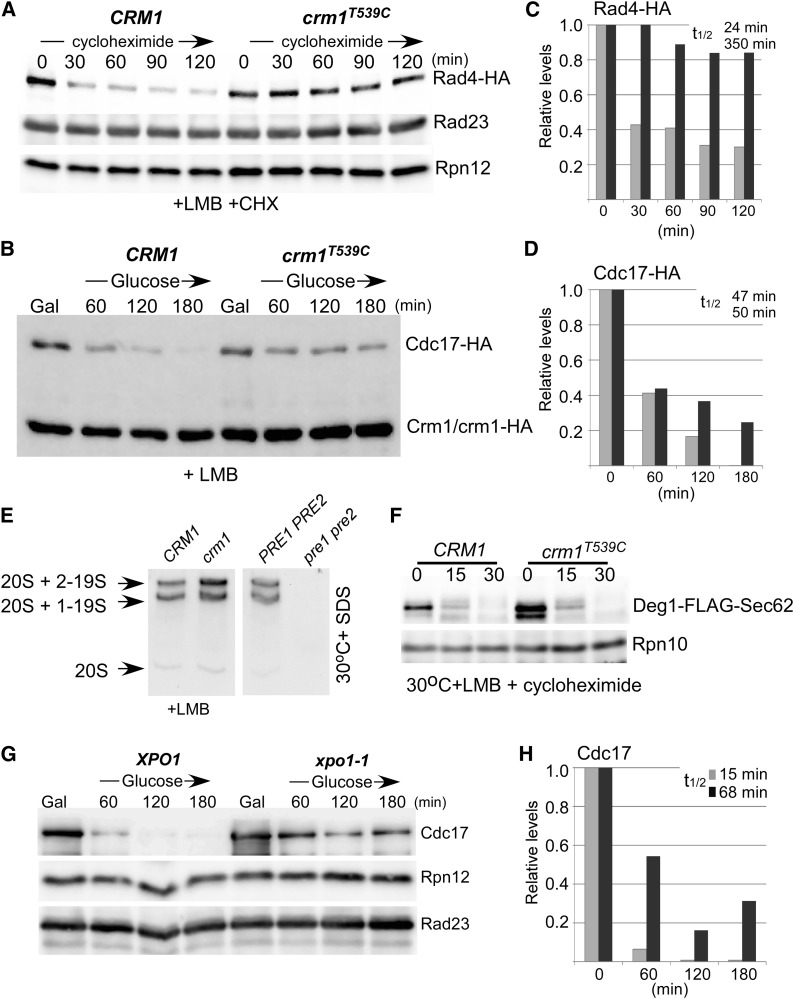
Stabilization of proteasome substrates in nuclear export mutants. (A and B) Epitope-tagged nuclear proteins (Rad4–HA and Cdc17–HA) were expressed in CRM1 and crm1T539C. LMB was added to actively growing cells at 30° and protein stability was determined after addition of cycloheximide to the growth medium. Both nuclear substrates were stabilized following inhibition of nuclear export (+ LMB in crm1T539C). The levels of internal controls (Rad23, Rpn12, Crm1–HA) are shown. (C and D) Rad4–HA and Cdc17–HA levels were quantified by densitometry. (E) Proteasome integrity and functionality was confirmed using a native in-gel fluorogenic assay, in which the hydrolysis of a chymotryptic substrate (LLVY–AMC) was examined. Proteasome peptidase activity was similar in CRM1 and crm1T539C. As expected, no chymotryptic activity was detected in the proteasome mutant pre1-1 pre2-2. (F) The turnover of Deg1–FLAG–Sec62 was unaffected in crm1T539C. (G) The stability of Cdc17–HA was also tested in the temperature-sensitive xpo1-1 export mutant. Cdc17–HA was expressed from the galactose-inducible PGAL1 promoter at 23° and protein stability was examined at the nonpermissive temperature (37°), after transfer to glucose medium. (H) The level of Cdc17–HA was measured by densitometry.
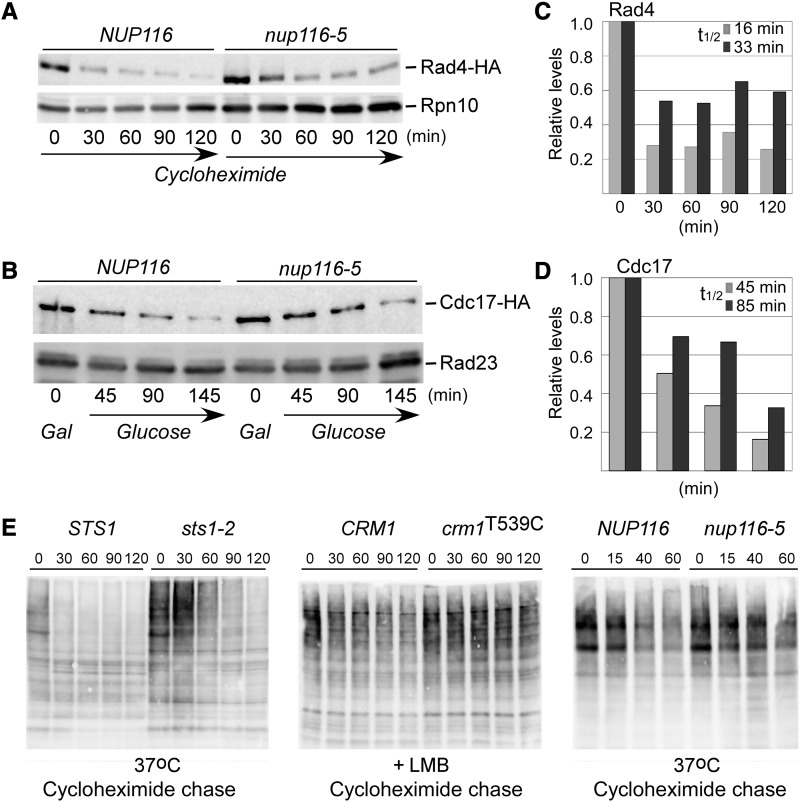
Nuclear substrates are stabilized when the nuclear pore is sealed. (A and B) Rad4–HA and Cdc17–HA were expressed in NUP116 and nup116-5. Cells were transferred from 23° to 37° and stabilization of Rad4–HA and Cdc17–HA was seen in nup116-5. Internal controls are shown (Rpn10, Rad23). (C and D) The levels of Rad4–HA and Cdc17–HA were quantified by densitometry, and values were adjusted to the 0-min sample for each protein. (E) The levels of overall multiubiquitinated proteins was determined in sts1-2 and nuclear transport mutants (crm1T539C and nup116-5). Protein extracts were prepared from STS1 and sts1-2 at 37° after the addition of cycloheximide (left). High levels of multi-Ub species (ranging from <10 to >250 kDa) accumulated in sts1-2. Similarly, protein extracts examined after the addition of LMB showed higher levels of multi-Ub proteins in crm1T539C (middle; + LMB) and in nup116-5 at the nonpermissive temperature (right; 37°).
References
-
- Cheong J. K., Gunaratnam L., Hsu S. I., 2008. CRM1-mediated nuclear export is required for 26 S proteasome-dependent degradation of the TRIP-Br2 proto-oncoprotein. J. Biol. Chem. 283: 11661–11676. - PubMed
-
- Enenkel C., Lehmann A., Kloetzel P. M., 1999. GFP-labelling of 26S proteasomes in living yeast: insight into proteasomal functions at the nuclear envelope/rough ER. Mol. Biol. Rep. 26: 131–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
