

Total body irradiation causes long-term mouse BM injury via induction of HSC premature senescence in an Ink4a- and Arf-independent manner
- PMID: 24622326
- PMCID: PMC4023419
- DOI: 10.1182/blood-2013-07-515619
Total body irradiation causes long-term mouse BM injury via induction of HSC premature senescence in an Ink4a- and Arf-independent manner
Abstract
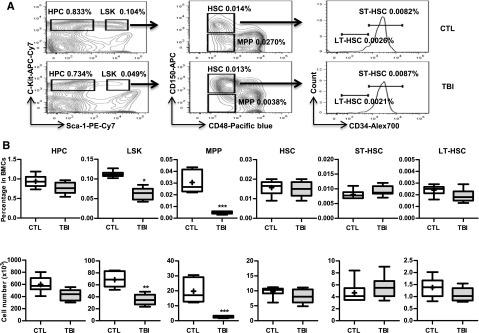
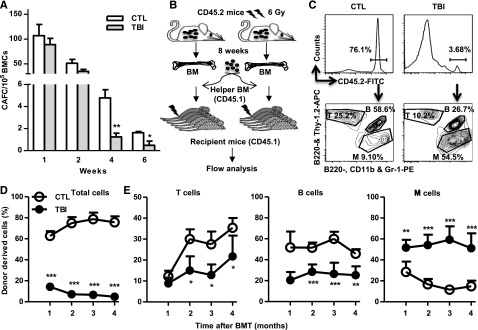
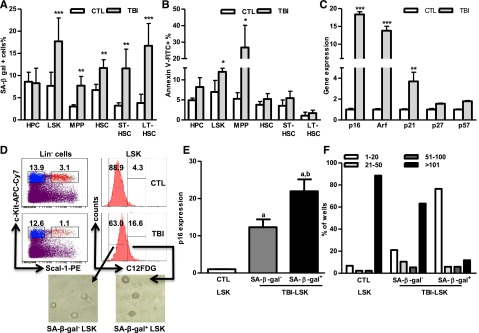
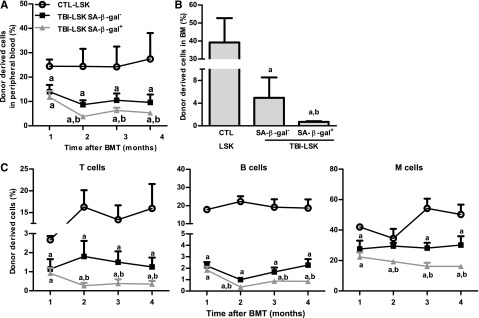
Exposure to total body irradiation (TBI) induces not only acute hematopoietic radiation syndrome but also long-term or residual bone marrow (BM) injury. This residual BM injury is mainly attributed to permanent damage to hematopoietic stem cells (HSCs), including impaired self-renewal, decreased long-term repopulating capacity, and myeloid skewing. These HSC defects were associated with significant increases in production of reactive oxygen species (ROS), expression of p16(Ink4a) (p16) and Arf mRNA, and senescence-associated β-galacotosidase (SA-β-gal) activity, but not with telomere shortening or increased apoptosis, suggesting that TBI induces residual BM injury via induction of HSC premature senescence. This suggestion is supported by the finding that SA-β-gal(+) HSC-enriched LSK cells showed more pronounced defects in clonogenic activity in vitro and long-term engraftment after transplantation than SA-β-gal(-) LSK cells isolated from irradiated mice. However, genetic deletion of p16 and/or Arf had no effect on TBI-induced residual BM suppression and HSC senescence, because HSCs from irradiated p16 and/or Arf knockout (KO) mice exhibited changes similar to those seen in HSCs from wild-type mice after exposure to TBI. These findings provide important new insights into the mechanism by which TBI causes long-term BM suppression (eg, via induction of premature senescence of HSCs in a p16-Arf-independent manner).
© 2014 by The American Society of Hematology.
Figures
Comment in
-
HSC senescence upon irradiation.Blood. 2014 May 15;123(20):3060-1. doi: 10.1182/blood-2014-03-563718. Blood. 2014. PMID: 24832936 Free PMC article.
References
-
- Mauch P, Constine L, Greenberger J, et al. Hematopoietic stem cell compartment: acute and late effects of radiation therapy and chemotherapy. Int J Radiat Oncol Biol Phys. 1995;31(5):1319–1339. - PubMed
-
- Testa NG, Hendry JH, Molineux G. Long-term bone marrow damage in experimental systems and in patients after radiation or chemotherapy. Anticancer Res. 1985;5(1):101–110. - PubMed
-
- Gardner RV, Begue R, McKinnon E. The effect of granulocyte-macrophage colony-stimulating factor (GM-CSF) on primitive hematopoietic stem cell (PHSC) function and numbers, after chemotherapy. Exp Hematol. 2001;29(9):1053–1059. - PubMed
-
- van Os R, Robinson S, Sheridan T, Mislow JM, Dawes D, Mauch PM. Granulocyte colony-stimulating factor enhances bone marrow stem cell damage caused by repeated administration of cytotoxic agents. Blood. 1998;92(6):1950–1956. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
