

Coordinated metabolic transitions during Drosophila embryogenesis and the onset of aerobic glycolysis
- PMID: 24622332
- PMCID: PMC4025483
- DOI: 10.1534/g3.114.010652
Coordinated metabolic transitions during Drosophila embryogenesis and the onset of aerobic glycolysis
Abstract
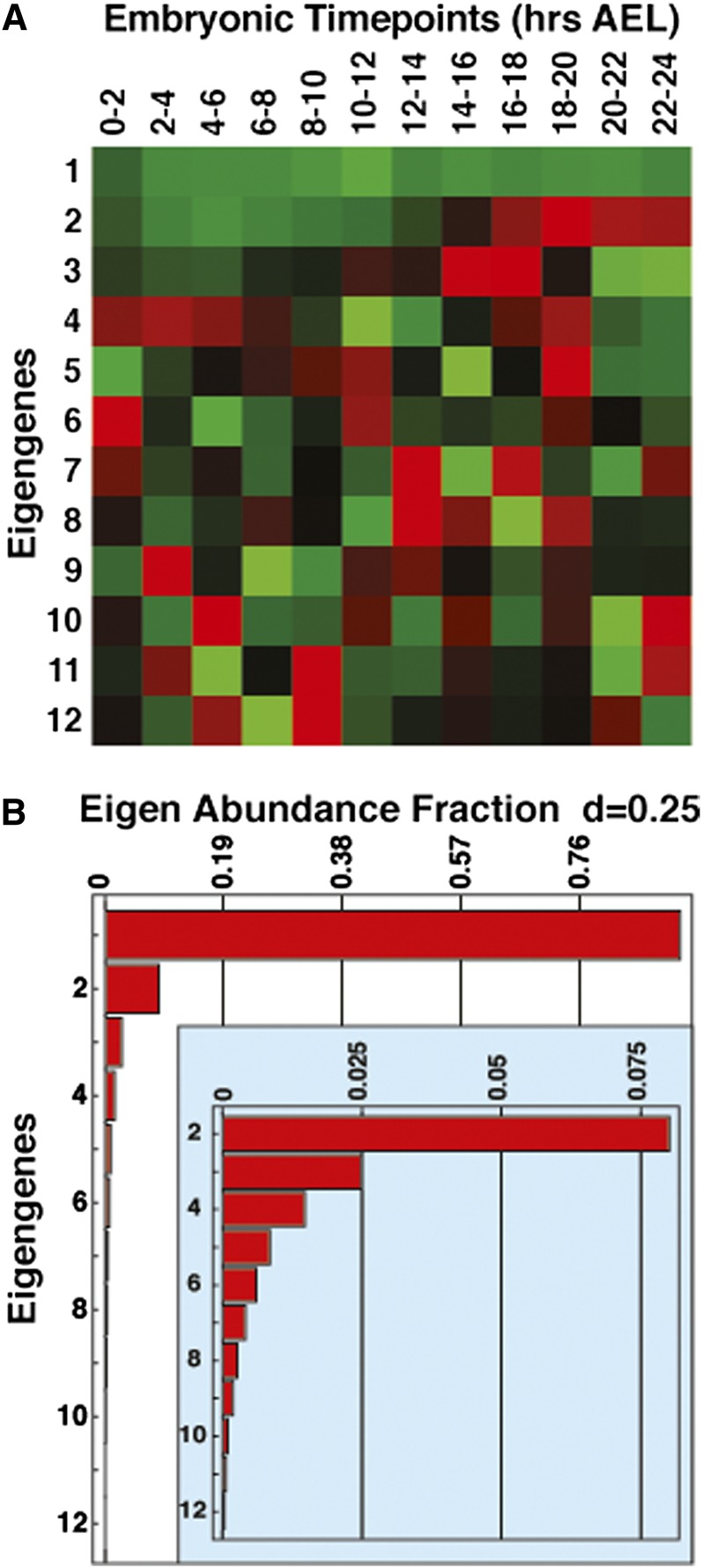
Rapidly proliferating cells such as cancer cells and embryonic stem cells rely on a specialized metabolic program known as aerobic glycolysis, which supports biomass production from carbohydrates. The fruit fly Drosophila melanogaster also utilizes aerobic glycolysis to support the rapid growth that occurs during larval development. Here we use singular value decomposition analysis of modENCODE RNA-seq data combined with GC-MS-based metabolomic analysis to analyze the changes in gene expression and metabolism that occur during Drosophila embryogenesis, spanning the onset of aerobic glycolysis. Unexpectedly, we find that the most common pattern of co-expressed genes in embryos includes the global switch to glycolytic gene expression that occurs midway through embryogenesis. In contrast to the canonical aerobic glycolytic pathway, however, which is accompanied by reduced mitochondrial oxidative metabolism, the expression of genes involved in the tricarboxylic cycle (TCA cycle) and the electron transport chain are also upregulated at this time. Mitochondrial activity, however, appears to be attenuated, as embryos exhibit a block in the TCA cycle that results in elevated levels of citrate, isocitrate, and α-ketoglutarate. We also find that genes involved in lipid breakdown and β-oxidation are upregulated prior to the transcriptional initiation of glycolysis, but are downregulated before the onset of larval development, revealing coordinated use of lipids and carbohydrates during development. These observations demonstrate the efficient use of nutrient stores to support embryonic development, define sequential metabolic transitions during this stage, and demonstrate striking similarities between the metabolic state of late-stage fly embryos and tumor cells.
Keywords: aerobic glycolysis; embryogenesis; metabolism; metabolomics.
Copyright © 2014 Tennessen et al.
Figures







References
-
- Backhaus B., Sulkowski E., Schlote F., 1984. A semi-synthetic, general-purpose medium for Drosophila melanogaster Drosophila Information Service. 60: 210–212.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous