

doi: 10.1128/JVI.00202-14.
Epub 2014 Mar 12.
Reinitiation after translation of two upstream open reading frames (ORF) governs expression of the ORF35-37 Kaposi's sarcoma-associated herpesvirus polycistronic mRNA
Affiliations
- PMID: 24623444
- PMCID: PMC4093840
- DOI: 10.1128/JVI.00202-14
Item in Clipboard
Reinitiation after translation of two upstream open reading frames (ORF) governs expression of the ORF35-37 Kaposi's sarcoma-associated herpesvirus polycistronic mRNA
J Virol.
2014 Jun.
Abstract
The Kaposi's sarcoma-associated herpesvirus (KSHV) ORF36 protein kinase is translated as a downstream gene from the ORF35-37 polycistronic mRNA via a unique mechanism involving short upstream open reading frames (uORFs) located in the 5' untranslated region. Here, we confirm that ORF35-37 is functionally dicistronic during infection and demonstrate that mutation of the dominant uORF restricts KSHV replication. Leaky scanning past the uORFs facilitates ORF35 expression, while a reinitiation mechanism after translation of the uORFs enables ORF36 expression.
Figures
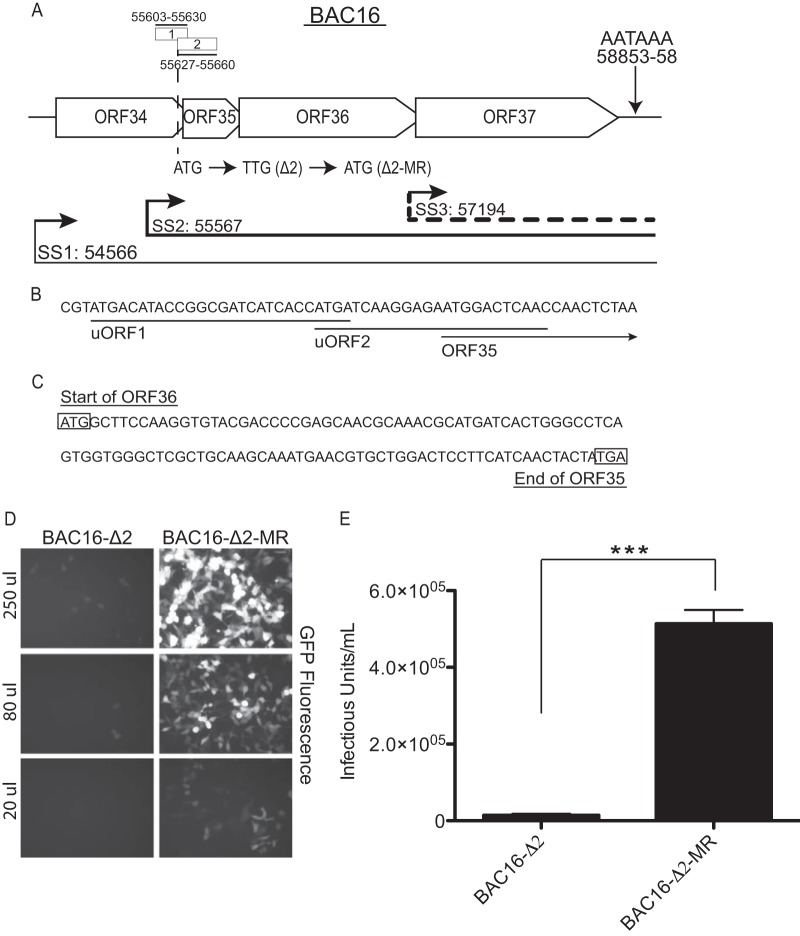
Disruption of uORF2 leads to a defect in the production of infectious viral particles. (A) A schematic presentation of the ORF34-37 genetic locus showing the ORF34-37, ORF35-37, and ORF37 mRNAs with thin, thick, and dashed lines, respectively. Start sites (SS) for each transcript and uORF1 and uORF2 genomic location are indicated according to the nucleotide position described by Russo et al. (30). The single poly(A) signal used by all four ORFs for transcription termination is shown. The nucleotide mutated to generate the uORF2 start codon mutant (ATG → TTG; Δ2) and the repaired marker rescue (TTG→ATG; Δ2-MR) is indicated. (B) Start and coding sequences for uORF1 and uORF2 and the N terminus of ORF35. (C) The region of overlap of sequence between ORF35 and ORF36. The ORF36 start codon and the ORF35 stop codon are boxed. (D and E) 293A cells (2 × 105 per well) were seeded 12 h prior to infection. At 48 h post-lytic reactivation with doxycycline, cell-free virus supernatant was harvested from the iSLK-PURO BAC16-Δ2 cell line or the Δ2-MR cell line, and then various dilutions were used to inoculate 293A cells via spinfection (2,000 × g for 45 min at 30°C). The cells were then incubated for 1 h, the inoculum was replaced with fresh media, and infection was allowed to proceed for 24 h prior to visualization by fluorescence microscopy (D) or analysis by flow cytometry (E). The percentage of GFP-positive 293A cells was detected using a FACSCanto II cell analyzer (BD Bioscience, San Jose, CA). Infectious unit (IU) values represent the number of GFP-positive cells in each well at the time of analysis. The experiment was performed in triplicate; error bars represent the standard deviations of the results of comparisons between replicates. Statistical significance was evaluated with a two-tailed unpaired t test (***, P < 0.0001).
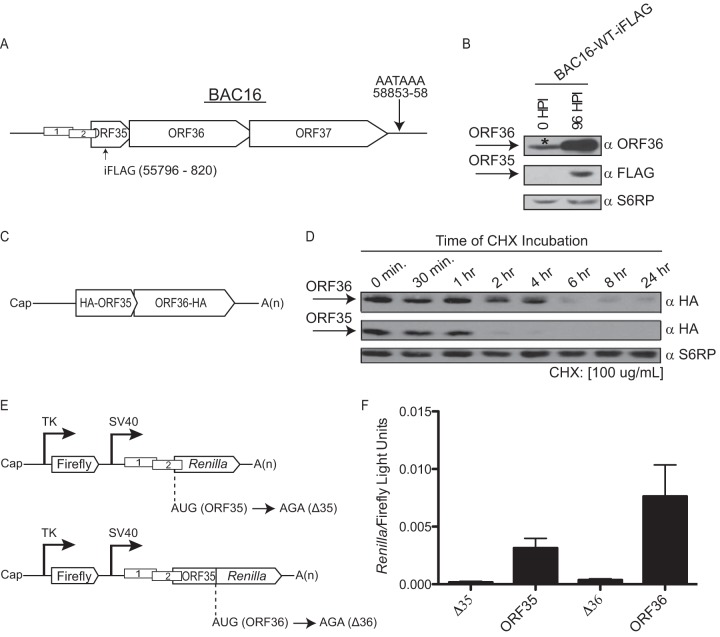
Analysis of the ratio of ORF35 expression to ORF36 expression during infection and transfection. (A) A schematic presentation of the ORF35-37 mRNA and the single poly(A) signal used by all four ORFs for transcription termination is shown. The position of the internal FLAG epitope tag (iFLAG) within ORF35 is indicated. (B) iSLK-PURO cells stably harboring the KSHV BAC16-iFLAG were either left untreated or lytically reactivated for 96 h. Protein lysates were subjected to Western blotting with antibodies against ORF36, FLAG-tagged ORF35, or the ribosomal protein S6RP (as a loading control). The asterisk (*) denotes a cross-reactive protein that comigrates with ORF36 in uninduced iSLK-PURO cells. HPI, hours postinfection; WT, wild type. (C) Schematic of the 5′ UTR hemagglutinin (HA)-ORF35-ORF36-HA plasmid, constructed as described in reference . (D) 293T cells were transfected with the 5′ UTR HA-ORF35-ORF36-HA construct. After 24 h, the cells were treated with 100 μg/ml of the translation elongation inhibitor cycloheximide (CHX), whereupon protein lysates were harvested at the indicated time points and immunoblotted with anti-HA antibodies or S6RP (as a loading control). (E) Schematic of the psiCHECK-2 (Invitrogen) dual-luciferase constructs that harbor firefly and Renilla luciferase under the independent control of the thymidine kinase (TK) and simian virus 40 (SV40) promoters, respectively. For the ORF35 construct, upstream of Renilla is the native 72-nucleotide (nt) 5′ UTR of the ORF35-37 mRNA containing uORF1 and uORF2 and the first 21 nt of ORF35. The Renilla AUG was mutated to AGA, causing its translation to initiate from the native ORF35 start codon and thus serve as a readout for ORF35 translation initiation. The ORF36 construct harbors the native 5′ UTR of the ORF35-37 mRNA, the uORF1 and uORF2 regulatory elements, and the first 352 nt of ORF35 followed by Renilla luciferase, which serves as a reporter for ORF36 translation initiation. The nucleotides mutated to disrupt AUGORF35 (Δ35) and AUGORF36 (Δ36) are shown. (F) The indicated constructs were transfected into 293T cells, and a dual-luciferase assay was performed 24 h posttransfection to determine the relative levels of Renilla luciferase activity and firefly luciferase activity. The experiment was performed in triplicate; error bars represent the standard deviations of the results of comparisons between replicates.
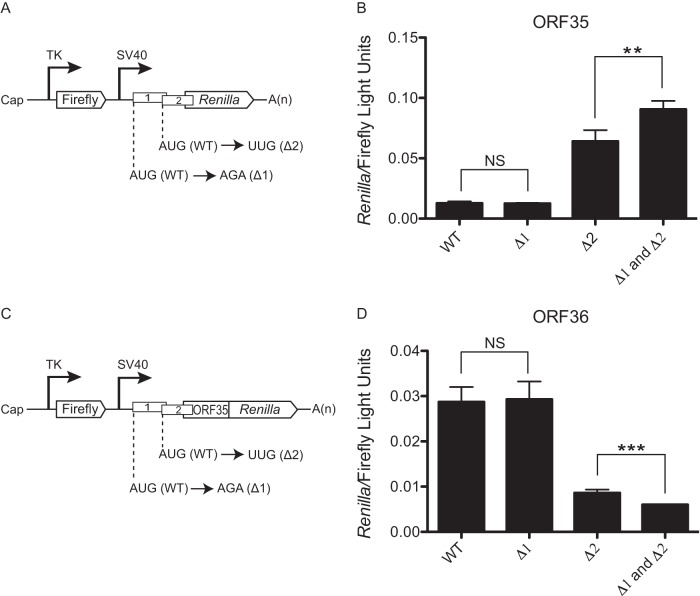
uORF1 and uORF2 govern expression of ORF35 and ORF36. (A and B) Schematics of the ORF35 (A) and ORF36 (C) reporter dual-luciferase constructs. The nucleotides mutated to disrupt AUGuORF1 (Δ1) and AUGuORF2 (Δ2) are shown. (B and D) The indicated constructs were transfected into 293T cells, and a dual-luciferase assay was performed 24 h posttransfection to determine the relative levels of firefly luciferase activity and Renilla luciferase activity. The experiment was performed in triplicate; error bars represent the standard deviations of the results of comparisons between replicates. Statistical significance was evaluated with a two-tailed unpaired t test (NS, P > 0.05; **, P < 0.001; ***, P < 0.0001).
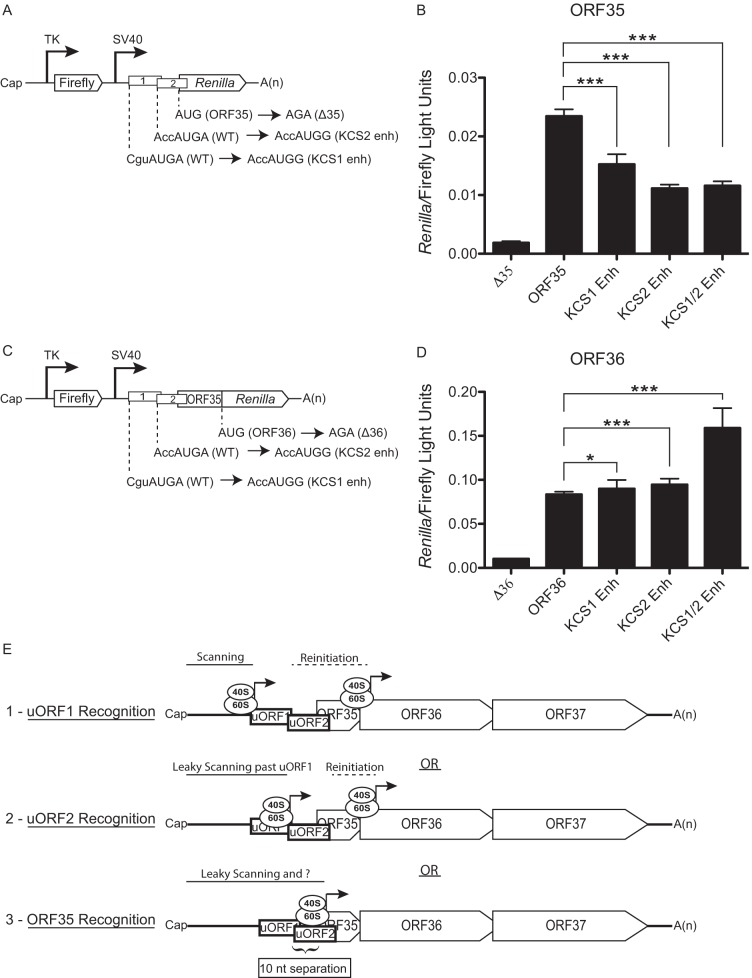
Leaky scanning past AUGuORF1 and AUGuORF2 facilitates ORF35 expression. (A and C) Diagram indicating the nucleotide mutations used to enhance the Kozak consensus sequence of the uORF1 (KCS1 enh) and/or the uORF2 (KCS2 enh) start codons within the ORF35 (A) and ORF36 (C) dual-luciferase reporter constructs. (B and D) The indicated constructs were transfected into 293T cells, and a dual-luciferase assay was performed 24 h posttransfection to determine the relative levels of firefly luciferase activity and Renilla luciferase activity. The experiment was performed in triplicate; error bars represent the standard deviations of the results of comparisons between replicates. Statistical significance was evaluated with a two-tailed unpaired t test (*, P < 0.05; **, P < 0.001; ***, P < 0.0001). (E) A model of the three translational programs governing expression from the ORF35-37 polycistronic mRNA. (1) Scanning ribosomes initiate translation at uORF1, terminate translation, continue to scan, and then reinitiate translation downstream of the ORF35 start codon. (2) Scanning ribosomes scan in a leaky manner past AUGuORF1 and initiate translation at AUGuORF2, terminate translation, continue to scan, and then reinitiate translation at the AUGORF36. (3) Scanning ribosomes scan in a leaky manner past AUGuORF1 and AUGuORF2 to initiate translation at AUGORF35. The 10-nt separation between AUGuORF2 and AUGORF35 is depicted.
References
-
- Martin DF, Kuppermann BD, Wolitz RA, Palestine AG, Li H, Robinson CA. 1999. Oral ganciclovir for patients with cytomegalovirus retinitis treated with a ganciclovir implant. Roche Ganciclovir Study Group. N. Engl. J. Med. 340:1063–1070 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA160556/CA/NCI NIH HHS/United States
- R01 CA082057/CA/NCI NIH HHS/United States
- CA31363/CA/NCI NIH HHS/United States
- R01 CA115284/CA/NCI NIH HHS/United States
- R01 AI073099/AI/NIAID NIH HHS/United States
- R01 CA136367/CA/NCI NIH HHS/United States
- R01 CA031363/CA/NCI NIH HHS/United States
- CA160556/CA/NCI NIH HHS/United States
- AI073099/AI/NIAID NIH HHS/United States
- R01 DE023926/DE/NIDCR NIH HHS/United States
- DE023926/DE/NIDCR NIH HHS/United States
- CA082057/CA/NCI NIH HHS/United States
- CA115284/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
