

Derivation of naive human embryonic stem cells
- PMID: 24623855
- PMCID: PMC3970494
- DOI: 10.1073/pnas.1319738111
Derivation of naive human embryonic stem cells
Abstract
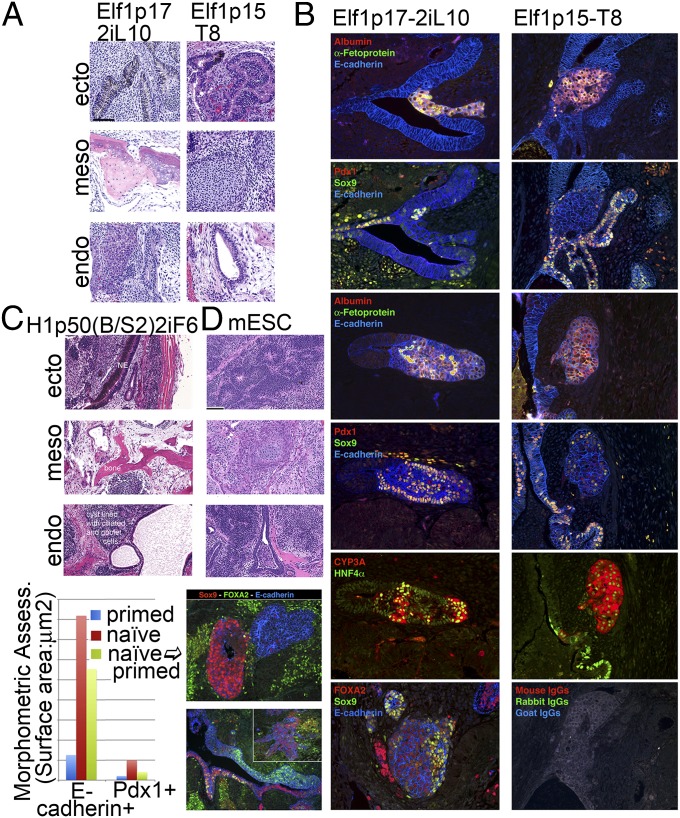
The naïve pluripotent state has been shown in mice to lead to broad and more robust developmental potential relative to primed mouse epiblast cells. The human naïve ES cell state has eluded derivation without the use of transgenes, and forced expression of OCT4, KLF4, and KLF2 allows maintenance of human cells in a naïve state [Hanna J, et al. (2010) Proc Natl Acad Sci USA 107(20):9222-9227]. We describe two routes to generate nontransgenic naïve human ES cells (hESCs). The first is by reverse toggling of preexisting primed hESC lines by preculture in the histone deacetylase inhibitors butyrate and suberoylanilide hydroxamic acid, followed by culture in MEK/ERK and GSK3 inhibitors (2i) with FGF2. The second route is by direct derivation from a human embryo in 2i with FGF2. We show that human naïve cells meet mouse criteria for the naïve state by growth characteristics, antibody labeling profile, gene expression, X-inactivation profile, mitochondrial morphology, microRNA profile and development in the context of teratomas. hESCs can exist in a naïve state without the need for transgenes. Direct derivation is an elusive, but attainable, process, leading to cells at the earliest stage of in vitro pluripotency described for humans. Reverse toggling of primed cells to naïve is efficient and reproducible.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
