

Fat-specific protein 27 (FSP27) interacts with adipose triglyceride lipase (ATGL) to regulate lipolysis and insulin sensitivity in human adipocytes
- PMID: 24627478
- PMCID: PMC4002109
- DOI: 10.1074/jbc.M113.539890
Fat-specific protein 27 (FSP27) interacts with adipose triglyceride lipase (ATGL) to regulate lipolysis and insulin sensitivity in human adipocytes
Abstract
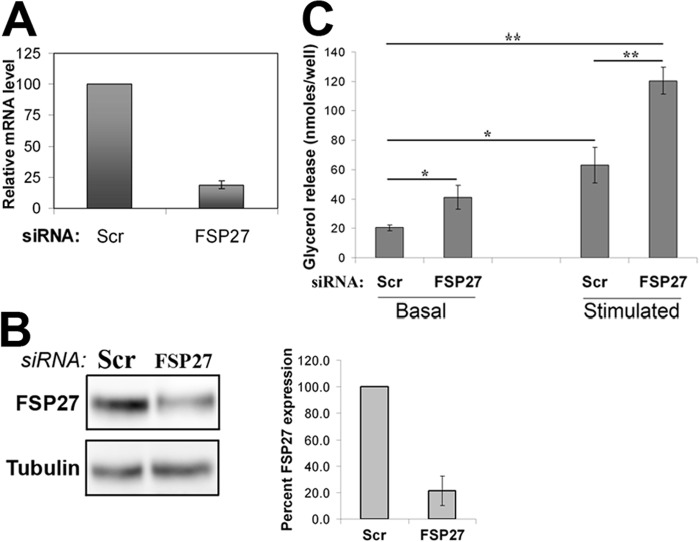
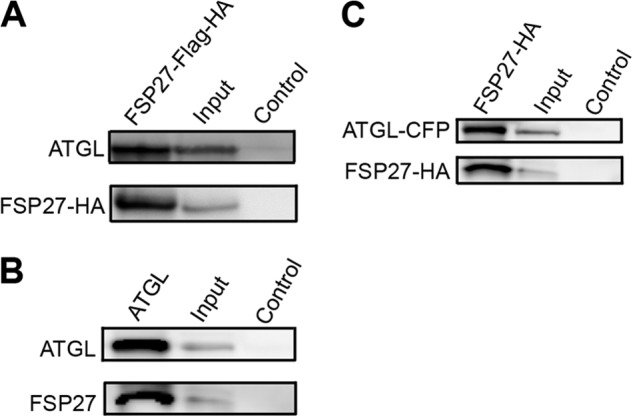
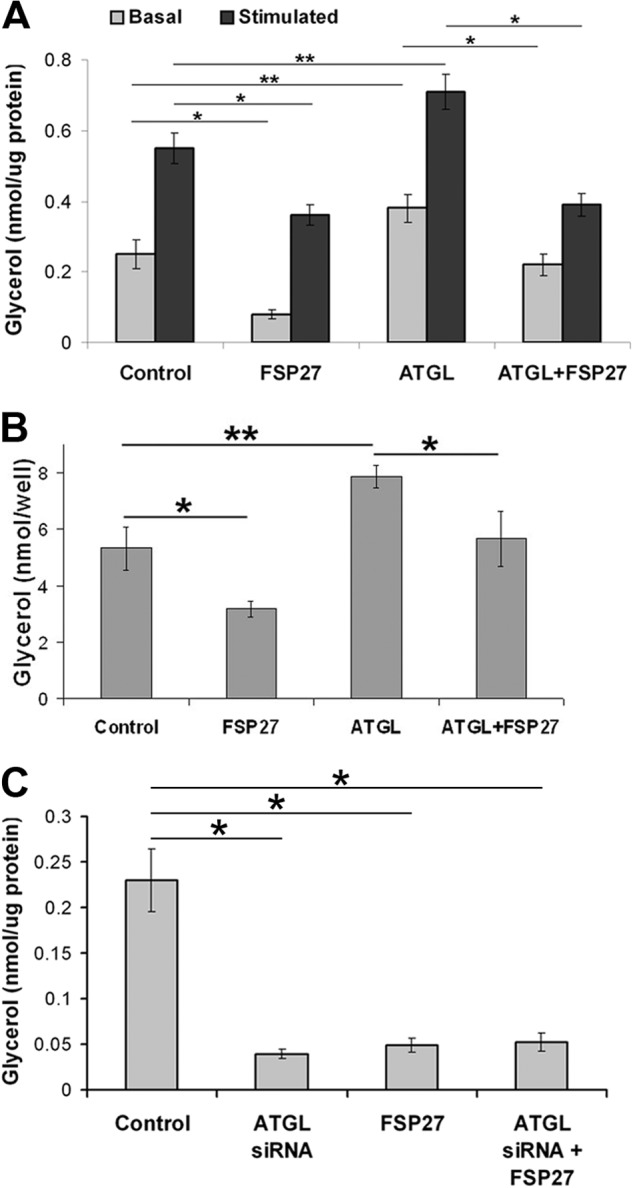
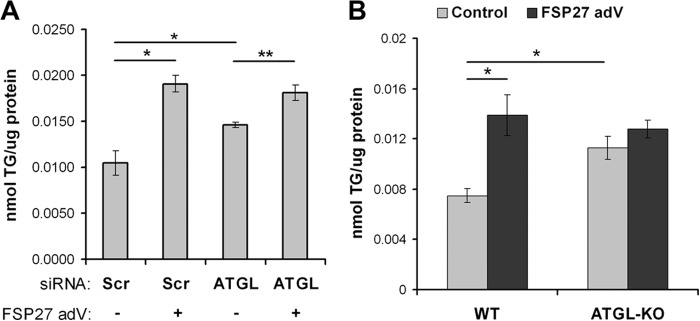
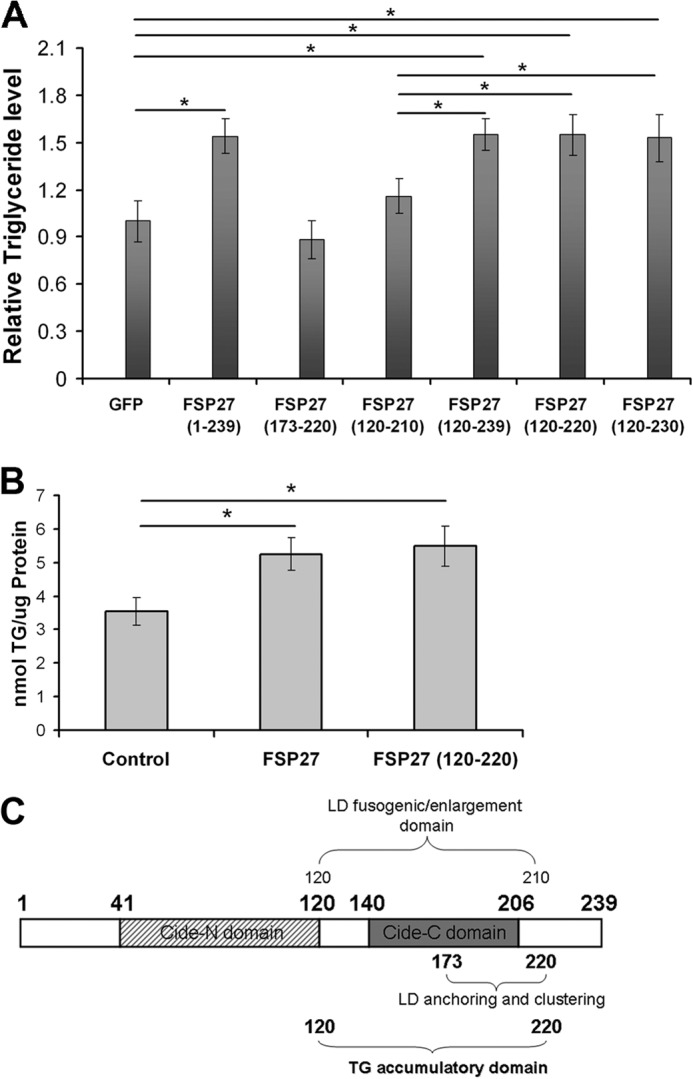
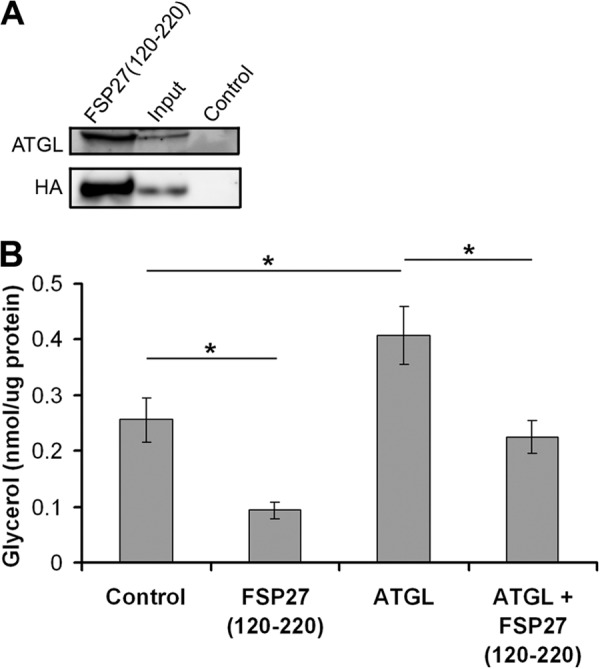
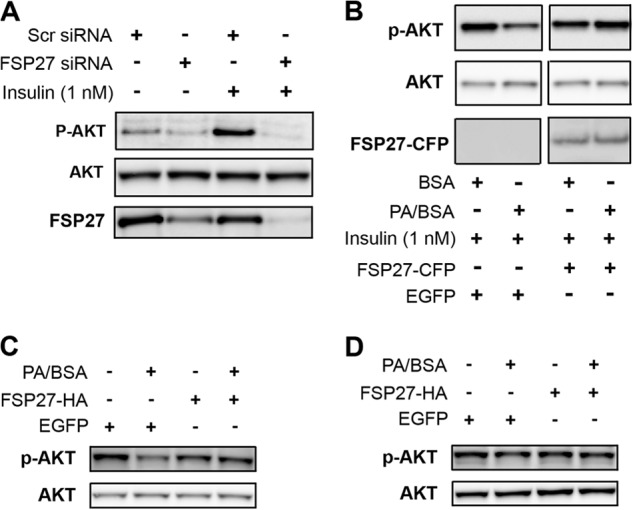
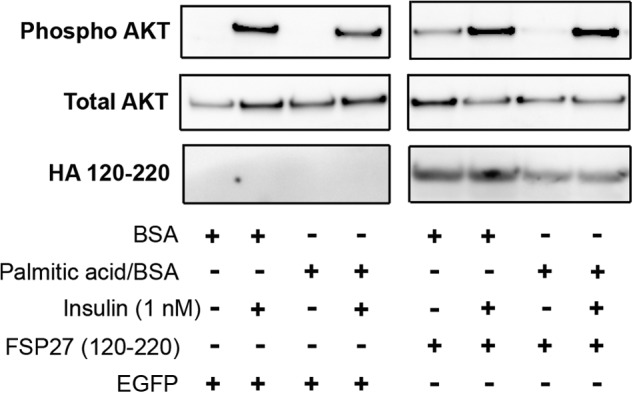
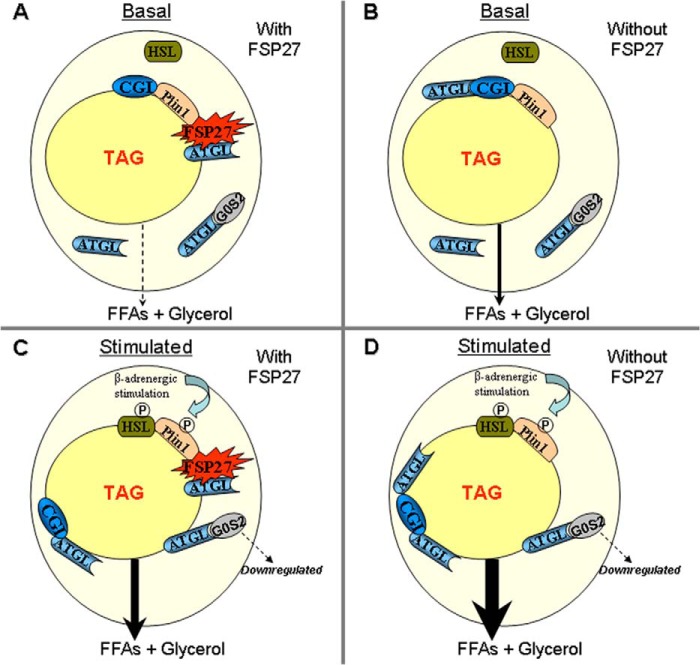
In adipocytes, lipolysis is a highly regulated process involving hormonal signals, lipid droplet-associated proteins, and lipases. The discovery of new lipid droplet-associated proteins added complexity to the current model of lipolysis. In this study, we used cultured human adipocytes to demonstrate that fat-specific protein 27 (FSP27), an abundantly expressed protein in adipocytes, regulates both basal and stimulated lipolysis by interacting with adipose triglyceride lipase (ATGL, also called desnutrin or PNPLA2). We identified a core domain of FSP27, amino acids 120-220, that interacts with ATGL to inhibit its lipolytic function and promote triglyceride storage. We also defined the role of FSP27 in free fatty acid-induced insulin resistance in adipocytes. FSP27 depletion in human adipocytes increased lipolysis and inhibited insulin signaling by decreasing AKT phosphorylation. However, reducing lipolysis by either depletion of ATGL or expression of exogenous full-length FSP27 or amino acids 120-220 protected human adipocytes against the adverse effects of free fatty acids on insulin signaling. In embryonic fibroblasts derived from ATGL KO mice, exogenous free fatty acids did not affect insulin sensitivity. Our results demonstrate a crucial role for FSP27-ATGL interactions in regulating lipolysis, triglyceride accumulation, and insulin signaling in human adipocytes.
Keywords: Adipocyte; Adipose Tissue Metabolism; Adipose Triglyceride Lipase; Diabetes; Diacylglycerol; Fatty Acid; Fatty Acid Metabolism; Insulin Resistance; Obesity.
Figures
References
-
- Unger R. H., Clark G. O., Scherer P. E., Orci L. (2010) Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim. Biophys. Acta 1801, 209–214 - PubMed
-
- Boden G. (1997) Role of fatty acids in the pathogenesis of insulin resistance and NIDDM. Diabetes 46, 3–10 - PubMed
-
- Dresner A., Laurent D., Marcucci M., Griffin M. E., Dufour S., Cline G. W., Slezak L. A., Andersen D. K., Hundal R. S., Rothman D. L., Petersen K. F., Shulman G. I. (1999) Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidylinositol 3-kinase activity. J. Clin. Invest. 103, 253–259 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F 3001/FWF_/Austrian Science Fund FWF/Austria
- 8KL2TR000158/TR/NCATS NIH HHS/United States
- Z 136/FWF_/Austrian Science Fund FWF/Austria
- UL1-TR000157/TR/NCATS NIH HHS/United States
- P 25193/FWF_/Austrian Science Fund FWF/Austria
- P30DK046200/DK/NIDDK NIH HHS/United States
- F 3002/FWF_/Austrian Science Fund FWF/Austria
- R56DK094815/DK/NIDDK NIH HHS/United States
- R56 DK094815/DK/NIDDK NIH HHS/United States
- KL2 TR000158/TR/NCATS NIH HHS/United States
- P30 DK046200/DK/NIDDK NIH HHS/United States
- UL1 TR000157/TR/NCATS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
