

Blood substitutes: why haven't we been more successful?
- PMID: 24630491
- PMCID: PMC4418436
- DOI: 10.1016/j.tibtech.2014.02.006
Blood substitutes: why haven't we been more successful?
Abstract
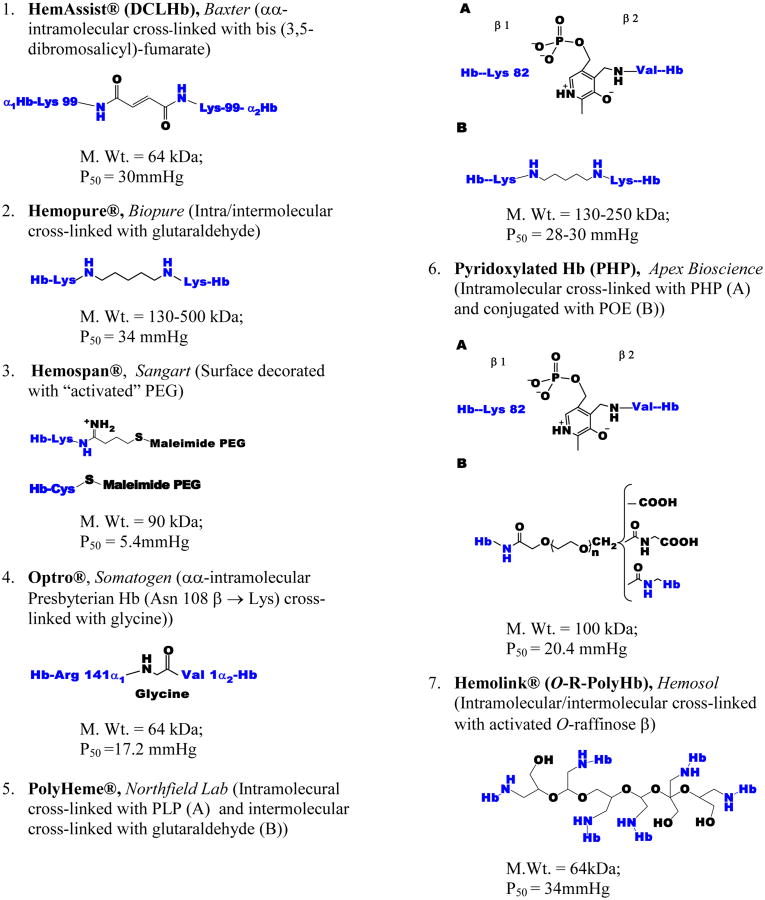
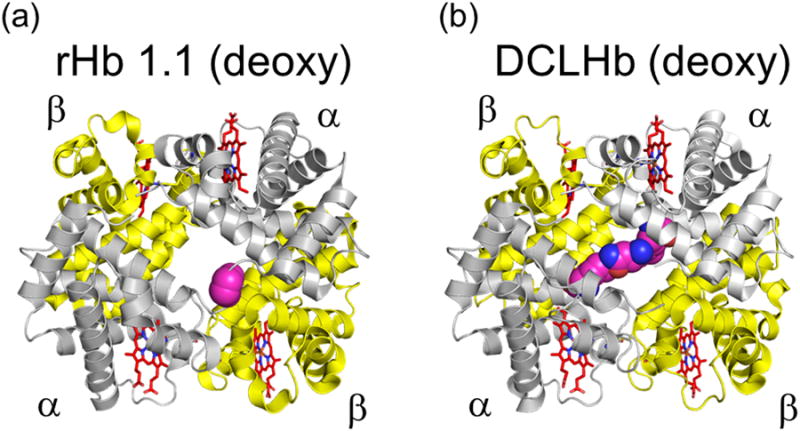
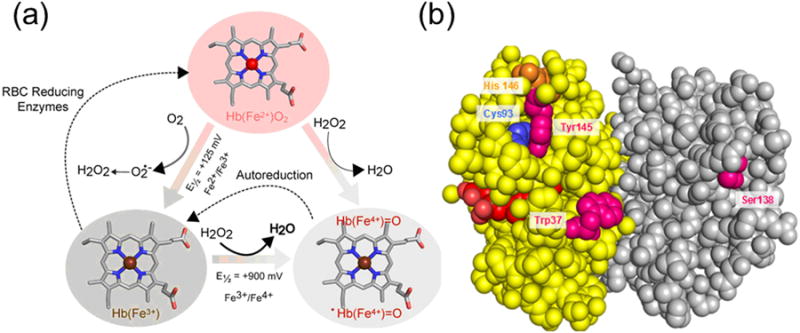
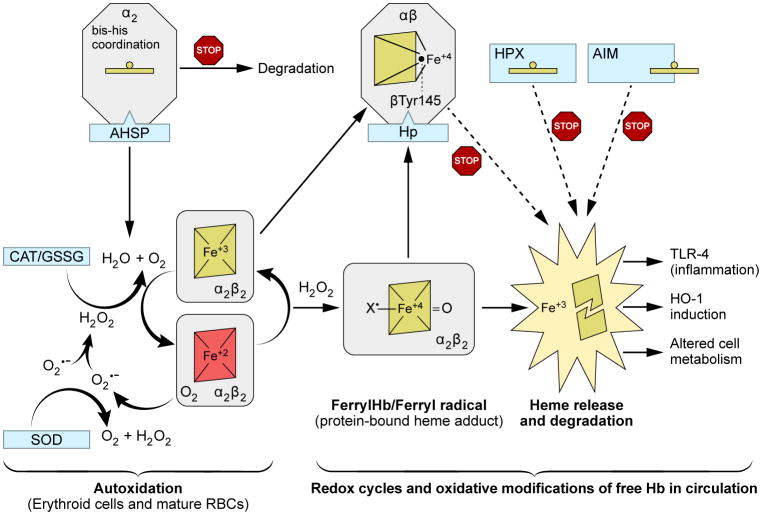
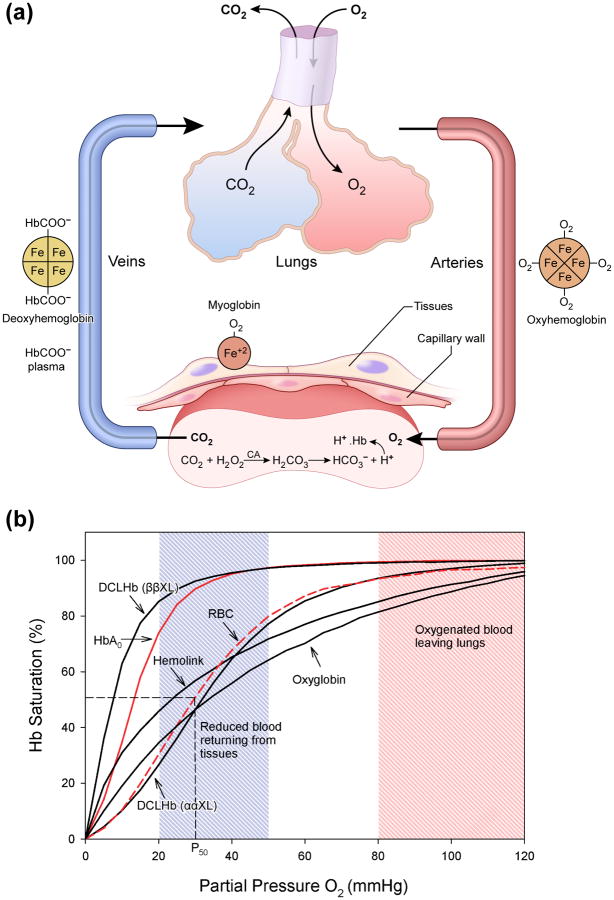
Persistent safety concerns have stalled the development of viable hemoglobin (Hb)-based oxygen carriers (HBOCs). HBOCs have several advantages over human blood, including availability, long-term storage, and lack of infectious risk. The basis of HBOC toxicity is poorly understood, however, several mechanisms have been suggested, including Hb extravasation across the blood vessel wall, scavenging of endothelial nitric oxide (NO), oversupply of oxygen, and heme-mediated oxidative side reactions. Although there are some in vitro and limited animal studies supporting these mechanisms, heme-mediated reactivity appears to provide an alternative path that can explain some of the observed pathophysiological changes. Moreover, recent mechanistic and animal studies support a role for globin and heme scavengers in controlling oxidative toxicity associated with Hb infusion.
Keywords: Blood substitutes; Heme; Hemoglobin; Oxidative toxicity.
Published by Elsevier Ltd.
Figures
References
-
- Kim HW, Greenburg AG. Artificial oxygen carriers as red blood cell substitutes: a selected review and current status. Art Organs. 2004;28:813–827. - PubMed
-
- Alayash AI. Setbacks in blood substitutes research and development: a biochemical perspective. Clin Lab Med. 2010;106:76–85. - PubMed
-
- Adamson JG, Moore C. Hemolink, an O-Raffinose crosslinked hemoglobin-based oxygen carrier. In: Chang TMS, editor. In blood substitutes, principles, methods, products and clinical trials. 1998. pp. 62–79.
-
- Dickerson RE, Geis I, editors. Hemoglobin:strcture, function, evolution, and pathophysiology. Benjamin/Cummings publishing Company; 1983.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
