

Growth conditions determine the DNF2 requirement for symbiosis
- PMID: 24632747
- PMCID: PMC3954807
- DOI: 10.1371/journal.pone.0091866
Growth conditions determine the DNF2 requirement for symbiosis
Abstract
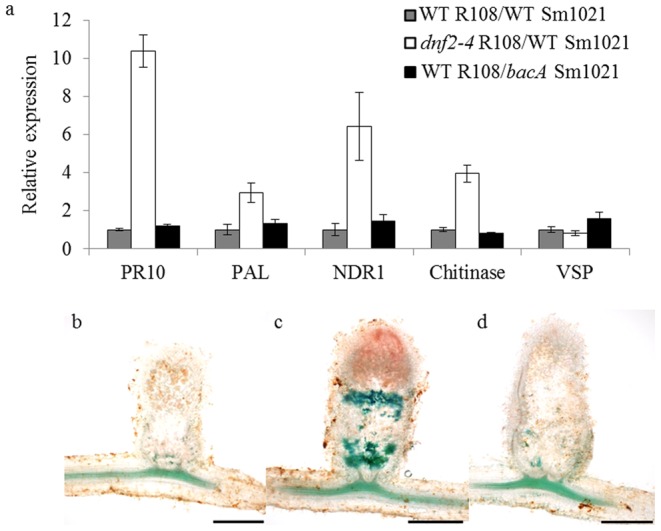
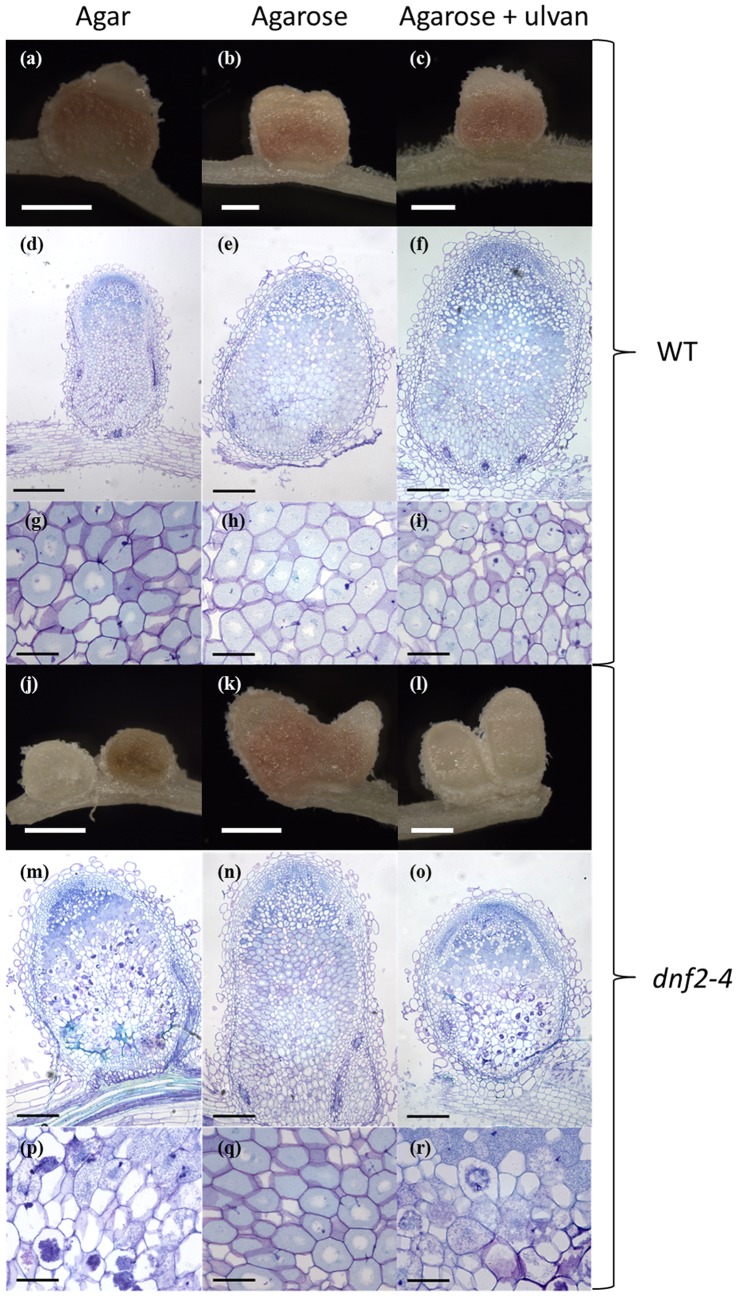
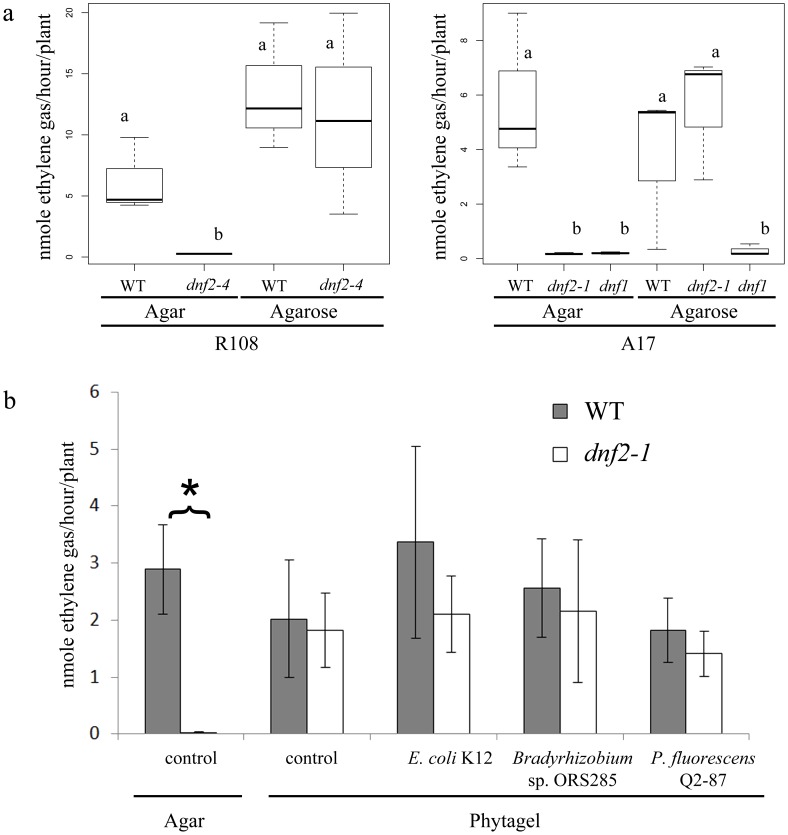
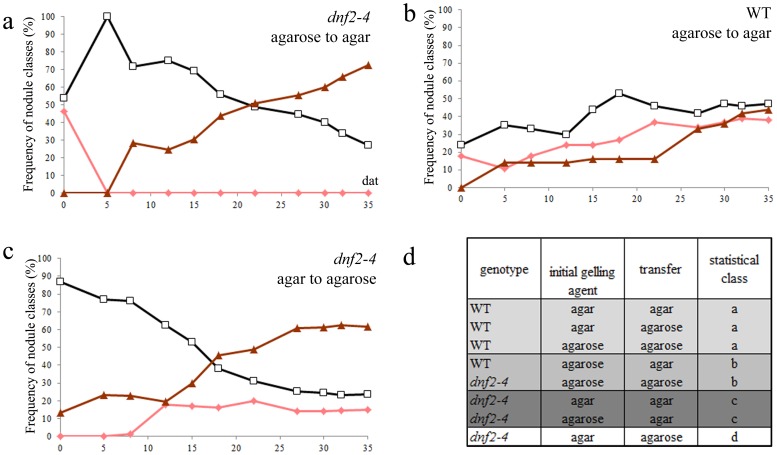
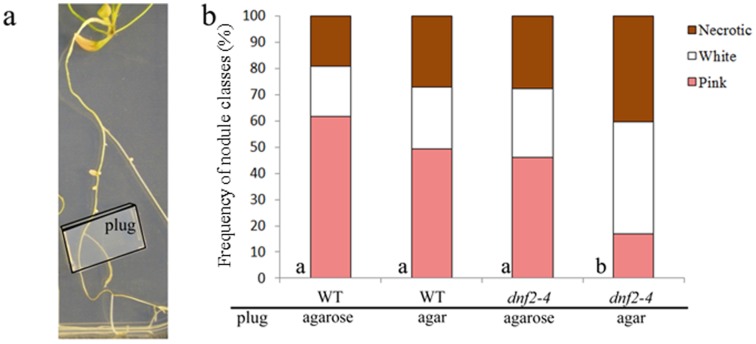
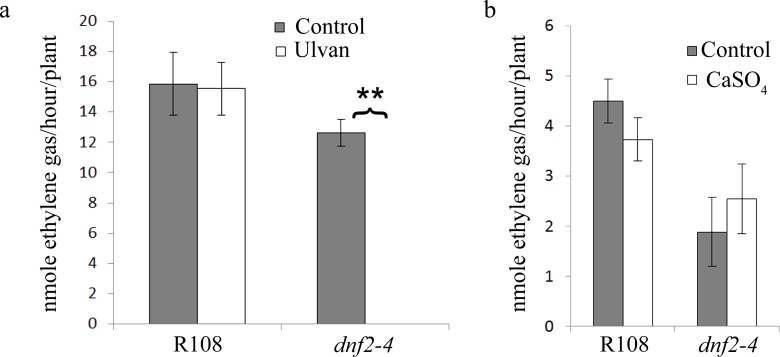
Rhizobia and legumes are able to interact in a symbiotic way leading to the development of root nodules. Within nodules, rhizobia fix nitrogen for the benefit of the plant. These interactions are efficient because spectacularly high densities of nitrogen fixing rhizobia are maintained in the plant cells. DNF2, a Medicago truncatula gene has been described as required for nitrogen fixation, bacteroid's persistence and to prevent defense-like reactions in the nodules. This manuscript shows that a Rhizobium mutant unable to differentiate is not sufficient to trigger defense-like reactions in this organ. Furthermore, we show that the requirement of DNF2 for effective symbiosis can be overcome by permissive growth conditions. The dnf2 knockout mutants grown in vitro on agarose or Phytagel as gelling agents are able to produce nodules fixing nitrogen with the same efficiency as the wild-type. However, when agarose medium is supplemented with the plant defense elicitor ulvan, the dnf2 mutant recovers the fix- phenotype. Together, our data show that plant growth conditions impact the gene requirement for symbiotic nitrogen fixation and suggest that they influence the symbiotic suppression of defense reactions in nodules.
Conflict of interest statement
Figures
References
-
- Puppo A, Groten K, Bastian F, Carzaniga R, Soussi M, et al. (2005) Legume nodule senescence: roles for redox and hormone signalling in the orchestration of the natural aging process. New Phytol 165: 683–701. - PubMed
-
- Charpentier M, Oldroyd G (2010) How close are we to nitrogen-fixing cereals? Curr Opin Plant Biol 13: 556–564. - PubMed
-
- Bourcy M, Brocard L, Pislariu CI, Cosson V, Mergaert P, et al. (2013) Medicago truncatula DNF2 is a PI-PLC-XD-containing protein required for bacteroid persistence and prevention of nodule early senescence and defense-like reactions. New Phytol 197: 1250–1261. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
